"Природа", №01-02, 1926 год, стр. 51-78.
"Если бы даже построение
Вегенера оказалось геофизи-
чески возможным, оно все
равно должно было бы рух-
нуть, если бы его не под-
твердили исторические фак-
ты: как правильно будет и об-
ратное: пусть протестует гео-
физик, историк будет продол-
жать работу над заинтересо-
вавшей его мыслью, будет
спокойно ждать, пока найдет
ей свое объяснение и геофи-
зик". Борисяк.
("Природа" 1922).
Изучение ареалов распространения систематических единиц растительного царства, а также и ареалов целых флор в их теперешнем объеме и прежнем распространении (по данным фитопалеонтологии), составляющее задачу генетической географии растений, наталкивается очень часто на невозможность объяснения происхождения этих ареалов действием современных нам факторов.
Это относится прежде всего к так называемым разорванным ареалам, где расстояние произрастания друг от друга представителей одной и той же систематической единицы — вида, рода или семейства растений, превосходит тот максимум расстояния, перенесение через которое может быть объяснено действием естественных факторов, как ветер, перенос птицами, морскими течениями и пр. Если отбросить возможность допущения одновременного возникновения совершенно однородных растений в различных местах земного шара (политопное развитие), то для объяснения таких разобщенных местонахождений нам необходимо будет прибегнуть к факторам, действовавшим в прежние геологические периоды.
Мы должны заранее допустить необходимость непрерывности распространения в прежнее время данной систематической единицы, ареал распространения которой охватывал, а часто и значительно превышал современные разобщенные местонахождения.
Эта разобщенность могла возникнуть под влиянием целого ряда факторов, к числу которых относятся иные климатические условия, при которых данная единица формировалась и распространялась, и вымирание ее на значительном протяжении этого ареала, вызванное изменением этих климатических условий. Далее такое же значение должно было иметь возникновение среди ареала непреодолимых преград, как горные массивы, пустыни и др., разобщившие ранее связанные местонахождения.
Но такого рода факторы являются совершенно недействительными для объяснения местонахождений разобщенных океанами или морями, если мы только допустим их неизменность, их перманентность. Нам нужно отказаться от последней и допустить необходимость связанности частей этих разорванных ареалов или непосредственной связанностью материков, на которых расположены отдельные части этих ареалов, или же через посредство ранее существовавших, а затем погрузившихся на дно океана пространств суши — как бы мостов, соединявших эти материки и обусловливавших прежнюю непрерывность ареалов.
Не меньшие трудности создает объяснение прежнего распространения флор в местностях, где в настоящее время они отсутствуют и где, при современных климатических условиях, их нахождение было бы совершенно невозможным. Объяснение такого резкого изменения климатических условий изменением положения полюсов, колебанием земной оси и др. еще далеко от того, чтобы быть вполне исчерпывающим.
В этом отношении каждая новая попытка объяснения происхождения материков и их расположения является чрезвычайно важной для биогеографии. И в то же время эта последняя дает геологии не менее ценную возможность проверки вновь созданной гипотезы.
С этой точки зрения представляют интерес попытки проверки на основании ботанико-географических данных теории Вегенера о происхождении материков.
В теории Вегенера1), которой уже не раз уделялось нашим журналом внимание2), есть для биогеографии два наиболее существенных момента. Это, во-первых, прежде существовавшая связанность всей суши в один материк, впоследствии расколовшийся на отдельные разошедшиеся друг от друга части, и во-вторых, изменение положения полюсов в связи с перемещением этой суши или самих полюсов, что создавало на ней изменения климатических условий и распределения климатических зон.
Связанность материков в один исходный континент существовала еще в триасе и только в юрском периоде начинается распадение его, выразившееся в намечающемся образовании двух меридиональных линий разлома. Эти линии разлома проходили между Европой-Африкой и Южной Америкой, с одной стороны, и между Африкой и Индией, с другой.
Отделение Южной Америки от Африки в меловом периоде делается все более и более заметным, хотя незначительная связь северо-восточного берега Ю. Америки и западного берега Африки, в его центральной части, существовала еще в третичном периоде, так что окончательное изолирование Ю. Америки и Африки произошло лишь после эоцена. Вместе с этим море, отделившее С. и Ю. Америки от Африки, делается все шире и шире, вследствие отхождения Америки к западу.
Связь Африки с Индией через Мадагаскар существовала еще в начале третичного периода и только в эоцене, вследствие движения Индии в направлении севера, эта связь порвалась.
Вместе с тем в юрском периоде происходит отделение Австралии от Индии-Цейлона, а также Антарктики от Южной Африки, которые сохраняя связь с Ю. Америкой двигались в юго-восточном направлении. В течение третичного периода происходит отделение Австралии от Антарктического материка, сохранявшего до четвертичного периода связь с Америкой; только в четвертичном периоде, вследствие движения последней на запад, произошел разрыв между Ю. Америкой и Антарктикой и все большее движение последней в направлении современного южного полюса.
В то же время, в течение ледникового периода, происходит отделение Гренландии от С. Америки и Европы, повлекшее за собой изолирование последних одна от другой.
Из сказанного мы можем сделать вывод, что сообщение Европы с С. Америкой существовало до четвертичного периода, Африки с Ю. Америкой до эоцена, Африки с Индией также до эоцена, Австралии с Индией до юрского периода, с Антарктикой и Ю. Америкой до середины третичного периода, и, наконец, Антарктики с Ю. Америкой до четвертичного периода. Соединение С. и Ю. Америк обусловлено не случайным соприкосновением, а существует с очень давних времен с той лишь разницей, что эта связь временами существовала лишь под водой, временами же соединявшие их участки суши выступали над поверхностью моря. Последнее имело место, вероятно, в силуре и девоне, затем с пермского периода до среднего триаса, затем опять в меловом периоде, и, наконец, с миоцена до наших дней.
Таков исторический процесс современного распределения материков, согласно Вегенеру. Положение последних по отношению к полюсам и друг к другу, нарушение связи между ними — не могло не отразиться на распределении растительности, и следы этого исторического процесса, несомненно, могут быть выявлены исторической географией растений.
География растений прежних геологических периодов является для нас во многих случаях еще далеко не объяснимой. Такой еще не разрешенной загадкой является распространение растений и целых флор в зонах земного шара, совершенно несоответствующих этим последним по своим современным климатическим условиям, а также однородность растительности, свидетельствующая об однородности климатических условий там, где в настоящее время имеется ясно выраженная смена климатических и растительных зон.
К числу таких загадок относится отсутствие признаков периодичности в росте растений каменноугольного периода. Эта периодичность у современных нам растений обусловливается сменой периода интенсивного вегетативного развития периодом покоя, соответствующему неблагоприятному в климатическом отношении времени года. Одним из характерных признаков такой периодичности являются, напр., у лиственных и хвойных растений спящие почки, остающиеся в закрытом состоянии в неблагоприятные периоды года и развивающиеся с наступлением благоприятствующих климатических условий. У растений каменноугольного периода, несмотря на наличие крупных деревьев, как напр, лепидодендроны, кордаиты, каламиты и др., такие почки совершенно не наблюдаются.
Другим, еще более характерным признаком периодичности являются годичные кольца утолщения древесины, характерные для растений, имеющих вторичное утолщение последней. У растений каменноугольного периода, на значительной части северного и отчасти южного полушария, колец утолщения не наблюдается. Это обстоятельство, а также и отсутствие спящих почек, свидетельствующих о беспрерывном и равномерном росте, могли иметь место лишь при ровном климате, лишенном периодической смены времен года.
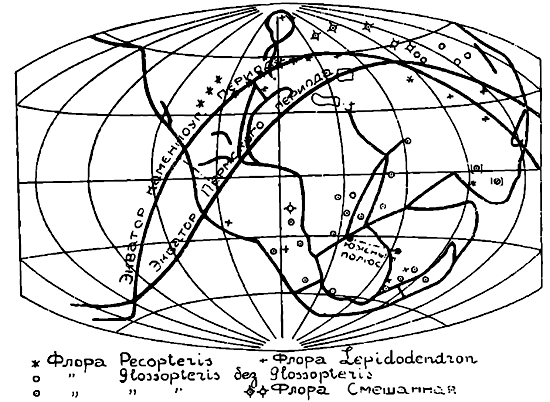
Такого рода климатические условия в настоящее время соответствуют экватору и характеризуют тропическую растительность. Между тем, растения каменноугольного периода, лишенные такой периодичности, произрастали на всем протяжении Европы.
Во флоре каменноугольного периода можно, согласно Потонье, различать элемент тропического происхождения, характеризующийся наличием папоротника Pecopteris, и элемент субтропический, характеризующийся наличием лепидодендронов, сигиллярий и др. В то время как первая — пекоптерисовая флора в каменноугольном периоде занимала вполне ограниченную, относительно узкую полосу, которая проходила через северную Америку, Европу и заканчивалась в восточной Азии, субтропическая — лепидодендровая флора лежала по обе стороны этой полосы и имела значительно большее распространение — она известна на севере до Шпицбергена, а на юге до южной Африки, т.-е. занимала в ширину интервал в 120°.
Другой, не менее трудно разрешимой загадкой, чем указанное отсутствие периодичности роста и широкое распространение тропических и субтропических флор вне современных тропиков и субтропиков, является произрастание деревьев, и притом субтропического характера, вблизи современных полюсов, напр., на Шпицбергене. При чем, что является совершенно непонятным, длительность полярной ночи видимо не оказывала на эту растительность никакого влияния.
Эти загадочные особенности в распространении и биологии ископаемых флор вызвали ряд попыток их объяснения. Так, делались допущения, что растения предшествующих геологических периодов обладали значительно большей, чем современные растения, способностью приспособления к различным климатам и были значительно менее чувствительны к теплу и холоду. Для объяснения несоответствия палеоклиматических особенностей, главным образом, равномерности климата на значительном протяжении земной поверхности, высказывались предположения, что однородность климата обусловливалась большим количеством внутреннего тепла земного шара и незначительной потерей этого тепла излучением, вследствие наличия покрова облаков, окружавших земной шар. Наконец, считали возможным допустить, что отсутствие колец утолщения есть особенность тогдашних растений и на основании их отсутствия нельзя делать никаких заключений, касающихся климата.
Эти положения опровергаются, с одной стороны, нахождением следов ледников в древнейших отложениях земли, а также тем, что уже в каменноугольном периоде однородность климата не была повсеместной, т. к. нам известны остатки древесины — с Фалькландских островов и из Австралии, имеющие кольца утолщения, с другой стороны, во флорах последующих периодов периодичность в росте растений все делается более и более распространенной и заметной. Это обстоятельство указывает на наличие климатических зон в предшествующие периоды жизни земли, но в то же время не оставляет сомнения в том, что положение земных материков в отношении этих зон было совсем иное. Вместе с тем, мы не можем допустить коренного отличия в физиологии и биологии растений прежних геологических периодов, и, даже наоборот, на основании данных фитопалеонтологии должны признать их полное тождество с современными. Согласно этим данным, ископаемые флоры должны были произрастать в климатических зонах, отвечающих физиологическим особенностям этих растений, которые в отношении требований к теплу и свету должны отвечать таковым и их современных потомков.
Все вышеизложенное заставляет нас допустить необходимость иного расположения полюсов в отношении материков. Из всех этих трудностей географии растений прежних геологических периодов выводит нас изложенная теория Вегенера, при свете которой климаты прежних геологических периодов получают совершенно иное и вполне нас удовлетворяющее объяснение.
Согласно реконструкциям Вегенера, мы должны допустить для каменноугольного и пермского периодов (см.карту 1) такое положение земной суши, бывшей еще в то время в связанном состоянии, при котором южный полюс находился на 25° южной широты и 25° восточной долготы, в пермском на 40—45° южной широты и 40° восточной долготы. Северный полюс в это время должен был лежать на 25° северной широты и 155° западной долготы, вследствие чего он со всех сторон был окружен океаном.
Такое положение полюсов должно было повлечь за собой и соответственное распределение климатических поясов. Экватор в каменноугольном периоде, согласно этой реконструкции, проходил через западную С. Америку, среднюю Европу и Каспийское море сильно выпуклой дугой к Зундскому Архипелагу. Средняя Европа, таким образом, находилась в полосе экваториальных дождей и в значительной степени была затоплена морем. Таким образом, каменноугольные залежи Средней Европы должны быть тропического происхождения, что и подтверждается образованием этих залежей флорой типа Pecopteris. Так разрешается вопрос о нахождении в каменноугольном периоде тропической флоры, лишенной периодичности в своем росте, в Средней Европе, а связанность материков объясняет однородность климатических условий на значительном протяжении последних. Вот эти-то перемещения полюсов, дают возможность нового объяснения и нового обоснования палеогеографии растений. Но обратимся теперь к современным флорам и их географическому распространению и посмотрим, насколько новая гипотеза Вегенера может разрешить те затруднения, которые возникают при изучении разорванных ареалов распространения современной нам растительности. Мы можем дать ответ на этот вопрос благодаря ряду проверок этой гипотезы, на основании изучения распространения отдельных групп растительного царства. Так, Сведелиус выполнил эту работу для некоторых тропических и субтропических водорослей, Кох — для цикадовых и хвойных, а Ирмшер — для покрытосемянных3).
Географическое распространение водорослей еще относительно очень мало известно. Это не касается морей и частей океанов, прилегающих к некоторым частям Европы, как Англия, Франция, Скандинавский полуостров, но в отношении больших океанов в целом, кроме разве Ледовитого океана, знакомство с распределением в них водорослей и причинами такого распределения еще очень ограничено. Помимо того, критическое изучение водорослей с использованием изучения их анатомического строения показало, что некоторые из них, раньше относившиеся к одному виду, очень часто не только должны быть отнесены к разным видам, но иногда и к разным родам. А это обстоятельство уже совершенно меняет взгляд на генезис данной океанической флоры. В этом отношении большой интерес представляет предпринятое Сведелиусом изучение флоры водорослей тропической части Индийского и Тихого океанов и прилегающей к Центральной Америке части Атлантического океана (Карибское море). Как известно флора водорослей этих двух океанов, в тропических районах Старого и Нового Света, является чрезвычайно однородной, несмотря на разобщенность этих двух частей ареалов Американским материком.
Предположение о связанности этих двух участков разорванного ареала, к югу от мыса Доброй Надежды на юге Африки и Огненной земли на юге Америки, исключается, т. к. общие физико-географические условия океанов в этих местах и наличие холодных течений, позволяющих обитанию здесь таких характерных для северных и умеренных морей водорослей, как ламинариевые, являются совершенно неподходящими для жизни тропических видов водорослей.
Наличие этого разорванного ареала уже раньше вызвало потребность в его объяснении, при чем было сделано допущение, что первоначально условия обитания в районе мыса Доброй Надежды и Огненной земли благоприятствовали тропическим видам, так что в то время флора указанных участков, разорванного сейчас ареала, соединялась к югу от Америки и Африки. Впоследствии, вследствие изменившихся климатических условий морей в районе этих последних, когда-то связанный ареал, лежащий по обе стороны Америки, оказался разорванным. Но такое объяснение, изменением ныне действующих факторов, является далеко не достаточным для разрешения указанного вопроса, тем более, что о таком изменении климатических условий этих частей Тихого и Атлантического океанов нам ничего не известно. Точно также, нет никакого основания для допущения возможности акклиматизации тропических видов водорослей в этих более холодных водах океана, благодаря которой могла бы образоваться связь разобщенных сейчас участков указанных ареалов. Эта незначительная пластичность водорослей была недавно отмечена Sauvageau, который показал, что, несмотря на однородность среды, в которой живут водоросли, несмотря на легкую возможность переноса их морскими течениями, пароходами и пр., морские водоросли очень редко натурализируются в местностях, удаленных от районов их обычного обитания.
В 1905 г., Сведелиус впервые выдвинул другое объяснение разобщенности тропических ареалов Атлантического, Индийского и Тихого океанов, основывающееся на исторических причинах. Согласно его предположению эти разобщенные части ареала в прежние эпохи неоднократно соединялись сообщением Тихого и Атлантического океанов на месте теперешнего Панамского перешейка. При этом последний раз такое соединение, хотя и в виде узкого канала, имело место гораздо позже, чем это раньше предполагалось, а именно в конце третичного периода (в плиоцене).
Эта точка зрения была подкреплена изучением водорослей этих же районов Börgesen'ом, который показал, что среди зеленых водорослей имеется гораздо больше разобщенных сейчас видов, чем среди бурых и багряных водорослей. Эти выводы важны тем, что зеленые водоросли включают в себя гораздо больше древних типов, чем бурые и багряные, среди которых мы находим большое количество так называемых критических видов несомненно более молодого происхождения. Если признать точку зрения Сведелиуса, то будет понятно отсутствие этих относительно недавно сформировавшихся видов, одновременно по обе стороны Центральной Америки, так как, очевидно, они возникли уже после прекращения связи Атлантического и Тихого океанов через Панамское соединение.
Ряд примеров достаточно ясно иллюстрирует точку зрения Сведелиуса о связи сейчас разорванных ареалов тропических видов водорослей через соединение Атлантического и Тихого океана в конце третичного периода, и служат для нее вполне достаточным основанием. Вместе с тем эти исследования, хотя и недостаточные для детальной проверки теории Вегенера, дают основание автору прийти к заключению, что "большая часть древних родов водорослей имела преимущественное распространение в Индийском и Тихом океанах, откуда она уже мигрировала в Атлантический океан. Только в одном или двух из изученных случаев миграция имела место в обратном направлении. Таким образом, флора водорослей Атлантического океана должна рассматриваться, как более молодая, чем Тихого и Индийского океанов. Это не противоречит теории Вегенера, согласно которой Атлантический океан имеет более позднее происхождение, чем Индийский и Тихий океаны".
Происхождение цикадовых в свете теории Вегенера было изучено Францом Кохом4). Цикадовые относятся к числу древних семейств, начало развития которых надо отнести к Пермскому или Триасовому периодам, т.-е. как раз к тому времени, когда по Вегенеру началось расхождение материков, вследствие чего пересмотр и изучение истории их возникновения и их распространения представляет особый интерес.
Цикадовые должны были непосредственно возникнуть из каменноугольной флоры в области тогдашнего экватора, т.-е. в районе теперешней средней и восточной Северной Америки, средней Европы и прилегающей части Азии, расположенной на той же широте, поскольку последняя не была покрыта мелкими водами моря. Таким образом, возникнув в конце каменноугольного периода в области т.н.каменноугольного экваториального кольца, цикадовые в последующие геологические периоды распространились к северу и югу, охватив значительные районы тогдашних субтропиков, как-то: север Европы и Сибири, Шпицберген, Гренландию, с одной стороны, и Средеземноморскую область и северную Африку, с другой.
Об этом широком распространении свидетельствуют многочисленные ископаемые остатки цикадовых, которые должны быть отнесены по меньшей мере к 34 родам и 278 видам. Этот расцвет цикадовых достиг апогея к началу мелового периода, после которого начинается их упадок и замена их хвойными и покрытосемянными.
История распространения цикадовых, согласно Коху, представляется в следующем виде. Группа цамиевых, Zamiасеае, уже в Триасе должна была проникнуть из Средней Европы в "северо-атлантическую сушу“ (см. карту реконструкции Вегенера для этого периода) и отсюда уже позже распространилась к югу в Среднюю Америку и в северную часть Южной Америки. Из этой группы два рода Dioon и Ceratozamia нe пошли дальше Мексики, род же Zamiа представлен не менее чем 30 видами в Бразилии, Перу и Гранаде, Гватемале, на Антильских островах и во Флориде; сюда же относится и род Microcycas, растущий на острове Кубе. Таким образом, мы ясно можем проследить постепенное движение этой группы цикадовых в триасе и юре из субтропической средней Европы и "северо-атлантической суши" к югу, сначала в Среднюю Америку, а затем в Южную Америку и прилегающие к ней острова, прежде чем произошло отделение обеих Америк одна от другой и отрыв островов от Южной Америки.
Другая группа Zamiaceae продвинулась к югу от экватора Пермского периода — в Африку, где она, следуя за постепенным потеплением климатических условий Африки, достигла через восточную Африку самой Капской земли. От этой группы сохранился, между прочим, род Encephаlartus. Из восточной же Африки близкие формы проникли через Мадагаскар и восточную Индию в Австралию, благодаря долженствовавшему существовать соединению этих материков, образовавших одно целое. В Австралии мы находим сильно развитой род Macrozamia, близкий к африканскому Encephalartus и американскому Zamiа.
Группа собственно цикадовых — Суcadaceae, представленная родом Сусаs, свидетельствует также о своем африканском происхождении. Хотя в самой Африке она не сохранилась, но ее первого представителя — Cycas Thouоrsii мы находим уже на Мадагаскаре и Каморских островах. В непосредственной близости к этому виду стоит Cycas circinalis из восточной Индии. Отсюда эта группа должна была распространиться и в составлявшей тогда одно целое Австралии. Это должно было иметь место не позднее триаса, т. к. уже в начале юры указанные соединения должны были прекратиться. Наконец, восточно-азиатские формы (Cycas siamеnsis и revoluta) достигли своего теперешнего местонахождения через восточную Индию, а не через Австралию, т. к. последняя придвинулась к Юго-восточной Азии в геологически более позднее время. Путь переселения цикадовых из восточной Африки в Австралию минует Антарктический континент, в противоположность таковому хвойных. Это обстоятельство объясняется тем, что цикадовые, как более древняя группа растений, могли уже в триасе по непрерывному соединению Африки, через Мадагаскар и Индию, проникнуть в Австралию, тогда как позже возникшие хвойные должны были для этого воспользоваться обходным путем через Антарктику.
Таким образом, цикадовые в своих путях расселения отличаются от хвойных тем, что они как теплолюбивые растения продвигались все время к югу, следуя за тем же направлением перемещения субтропических зон. Во время же вторичного перемещения полюса к северу — в эоцене, они уже не были в состоянии следовать за этим новым изменением климатических условий, что повлекло за собой начало их постепенного вымирания. Произведенным исследованием, Кох считает, что он показал, что теперешнее распространение цикадовых в далеко разобщенных тропических и субтропических странах и распространение их ископаемых предков в теперешних умеренных зонах дождей и в частности в теперешних полярных странах, может быть удовлетворительно объяснено лишь на основе Вегенеровской теории перемещения полюсов и континентов.
Начало развития хвойных, если не считать кордаитовых, появляющихся уже в девоне, надо отнести к пермскому периоду. Если допустить, согласно Вегенеру, передвижение полюсов с запада на восток, то, как мы видим, полюс приходился на самом континенте в пермо-карбоновом периоде на южном, а в дилювии на северном полушарии. Противоположный полюс в это время должен был быть расположен каждый раз среди океана. Вследствие этого кажущаяся однородность климата северного полушария, как уже указано, не исключает наличия климатических зон, которые существовали, как и сейчас, но положение которых в отношении материков, соответственно другому расположению полюсов, было иное. Таким образом, как и сейчас, за экваториальной зоной тропических деревьев следовала более сухая субтропическая зона пустынь, за ней умеренная зона дождей и, наконец, полярная зона.
Первоначальный центр развития хвойных мы должны искать в умеренной зоне дождей, так как в них мы должны видеть первую группу растений, приспособившуюся к все нароставшему понижению температуры и необходимости уменьшения испарения воды. Вследствие этого продвижение хвойных и их постепенное развитие в течение ряда геологических периодов должно было следовать продвижению этой климатической зоны, которая в свою очередь изменяла свое положение вследствие перемещения полюсов. Поэтому понятно, что мы можем ожидать наличия (и на самом деле это имеет место) двух центров развития хвойных — одного в северном, другого в южном полушариях.
Эти обе группы хвойных, несмотря на современное совместное произрастание части видов, все же очень резко различаются и легко ограничиваются по морфологическим признакам и особенностям географического распространения (карта 2).
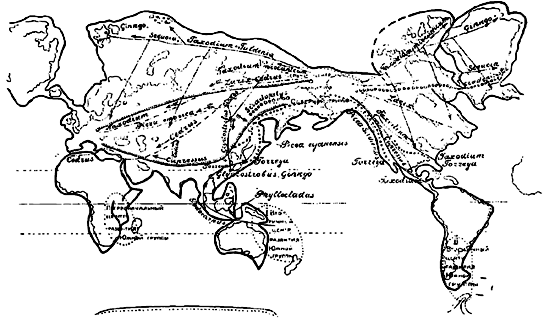
Представители южной группы еще в юрском, самое позднее в меловом периоде, продвинулись из Африки и Мадагаскара в восточную часть Антарктики, еще в то время находившуюся с ними в соединении. Отсюда путь хвойных лежит в Австралию, где они нашли благоприятные условия для своего развития, так что мы можем говорить здесь о вторичном, Австралийском центре развития южной группы хвойных. Действительно, в то время как Африканский центр — в восточной Африке, в Капской Земле и на Мадагаскаре — характеризуют немногие виды подокарпусов и кипарисовых, хвойные в Австралии и на прилегающих островах, в то время с ней связанных, достигают поразительного развития. Отсюда представители этой группы проникли в западную часть Антарктики, а затем, быть может, лишь в дилювии, благодаря существовавшей еще связи, в южную оконечность Ю. Америки. Эти последние виды уцелели до наших дней, в то время как сейчас их Антарктическая родина, вследствие изменения своего положения и положения полюса, погребена под толщей льда.
Южная группа в дальнейшем, после распадения южного континента на ряд материков, почти не имела дальнейшего развития. Только роды Araucaria, Dacrydium и в особенности Podocarpus, при дальнейшем распространении, образовали значительное количество форм. Вследствие этого у представителей этой группы мы находим ряд примитивных признаков и эндемичных реликтовых форм, которые могли сохраниться на оказавшихся изолированными участках суши, превратившихся в острова, подобно тому как здесь же только и сохранился ряд вымирающих, реликтовых животных. Вследствие этого понятно, что среди видов южной группы нет почти ни одного морозоустойчивого вида. Все они никогда не подвергались действию холода и поэтому для их существования необходимы океанические, теплые климатические условия. Только одна Araucaria imbricata, хорошо растущая у нас на Кавказском и Крымском побережье Черного моря, выдерживает зиму Германии. В пермском периоде северный полюс, согласно Вегенеру, приближался к Северной Америке и Беринговой Земле; вследствие этого в тогдашней умеренной зоне северного полушария мы должны искать первоначальный центр развития северной группы хвойных. Здесь, в месте будущего Берингова пролива, в то время существовали климатические условия, благоприятствующие развитию растений, приспособленных к прохладному и дождливому климату.
Пермокарбоновое оледенение должно было иметь место в северной части Тихого океана, но в юрском периоде эта зона оледенения должна была достигнуть широты современного Берингова пролива. Вследствие этого возникшие здесь, сейчас очень древние, виды продвинулись отсюда в Гренландию и Шпицберген, а также вдоль берегов Тихого океана: на юго-востоке — до Китая и Японии, на юго-западе — до Калифорнии и Флориды.
Эти древние виды отчасти сохранились и до настоящего времени и часто расположены почти симметрично по обе стороны Тихого океана.
В то время, как мы видели, часть хвойных ушла от надвигающегося полюса в район современной полярной области, где впоследствии и погибла, другая их часть распространилась от побережья Тихого океана на Запад и на Восток.
К этому времени относится сильное развитие хвойных северного происхождения, примером чего могут служить пихтовые. Этому способствовала большая протяженность и связанность континентов северного полушария в противоположность расчлененности их в южном полушарии.
За дальнейшим перемещением северного полюса следовало оледенение соответствующей зоны вокруг него, уничтожавшее в этой зоне всякую жизнь. В плиоцене полюс находился почти там же, где и в данный момент, так что вся растительность, покрывавшая современную полярную область, была уничтожена. Дальнейшее продвижение полюса в направлении С. Америки вызвало оледенение северной части последней и необходимость для растительности искать от него убежища на юге материка, что могло совершиться беспрепятственно вследствие отсутствия горных хребтов, которые могли бы преградить это движение к югу. Последнее имело место в Европе, когда, при перемещении полюса в сторону последней, растительность, спасавшаяся в южном направлении, была остановлена преградившими ей путь Альпами и почти вся уничтожена. Этим объясняется бедность видами хвойных Европы.
Итак, данные изучения распространения хвойных дают основание Коху считать, что распространение современных и ископаемых хвойных требует допущения перемещения полюсов в направлении вполне соответствующем теории перемещения материков Вегенера и становится на ее основе понятным.
Проверка теории перемещения материков на основе изучения покрытосемянных, произведенная Ирмшером, показала, что анализ ареалов распространения семейств последних дает возможность наметить в них некоторые общие черты. Эти данные позволяют нам установить некоторую закономерность в распространении семейств покрытосемянных, и прежде всего нельзя не отметить тот интересный факт, что из 289 семейств в Америке, Европе, Африке и Азии встречается 139 семейств, тогда как остальные 150 семейств распространены по всему земному шару. Только 33 семейства ограничены в своем распространении Новым Светом и 51 семейство — Старым Светом. Остальные 205 семейств общи и тому и другому. Только одно семейство свойственно лишь одному из пяти материков.
Закономерность характера ареалов этих семейств выражается, во-первых, в их симметричности, заключающейся в том, что центры преобладания развития образующих родов расположены таким образом, что по меридиану, проходящему через Европу—Африку, они или совсем отсутствуют (карта 3) или же представлены лишь очень незначительно, что связано часто с таким же незначительным распространением и в Австралии. Этот пропуск в ареале может быть объяснен лишь какими-то процессами в прежние геологические эпохи, нарушившими его целость. Вторая закономерность может быть установлена в виде той зональности, которую можно констатировать в строении разобщенных ареалов. Эта зональность выражается в том, что части разорванных ареалов располагаются в горизонтальном, а не меридиональном направлении, т. ч. мы их встречаем или в одной из умеренных зон, или в тропической зоне, или же в обеих умеренных зонах (биполярные ареалы).
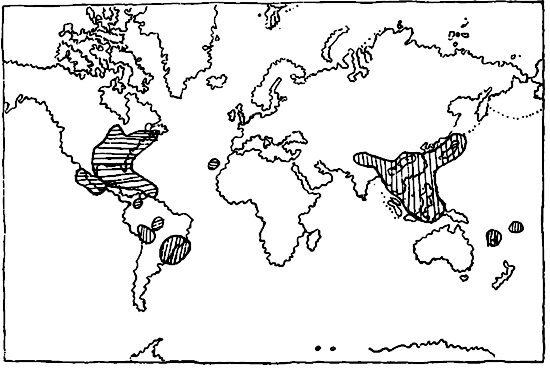
Третья законность может быть установлена в связанности ареалов распространения родов с определенными материками, при чем это выражается в очень незначительной связанности Америки с Европой—Африкой или в полном отсутствии таковой, и, наоборот, в значительном количестве ареалов, охватывающих Европу—Африку, Азию и Австралию. Эта связь ареалов отражает современное распределение материков — обстоятельство для дальнейших заключений весьма важное. Для уяснения современных ареалов покрытосемянных является весьма важным сравнение с теми же ареалами, но дополненными ископаемыми остатками тех же или близких родов и видов. Большинство таких ископаемых известно из северного полушария и преимущественно из С. Америки, Полярной области и Европы. При этом оказывается, что большинство форм в своем распространении было ограничено зоной — С. Америка—Европа, причем часть их — виды умеренного пояса и субтропиков — доходили до Полярной области, тропические же формы найдены б. ч. в средней и южной С. Америке, в Европе, а также в северной части Африки.
Данные такого сравнения указывают на три для нас очень важных факта. 1. Большое число разорванных ареалов, имеющие своих представителей в данное время лишь в Америке и Азии, имели их раньше в Европе и таким образом связывали сейчас разобщенные части ареала (карта 4). 2. Многие формы встречались раньше севернее, чем в настоящее время, т. е., иначе говоря, многие тропические семейства обитали в теперешних умеренных широтах Америки, а затем уже значительно позже в Европе и Азии (карта 5). На основании последних данных мы не можем не сделать вывода, что эти роды должны были возникнуть в Америке и затем уже отсюда проникли на восток.
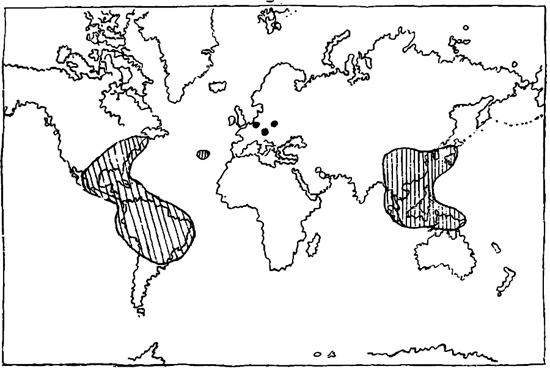
Сейчас отмеченное обстоятельство, что ископаемые местонахождения многих видов и родов находятся в зонах, в данное время по своим температурным условиям неподходящим для их произрастания, и вместе с тем установленная нами зональность требует тех же климатических условий и для ископаемых остатков, заполняющих разорванные ареалы. Если же эти ископаемые остатки находятся нами вне той же климатической зоны, т. к., принимая постоянство физиологических особенностей растений, мы должны допустить: или, что в то время эти климатические зоны имели большее широтное притяжение; или же, что они, сохраняя свои широтные размеры, подверглись передвижению с севера на юг. Разрешение этого вопроса мы находим в том обстоятельстве, что ископаемые остатки были находимы севернее произрастания их теперешних представителей лишь в Европе и отчасти Америке, но никогда на той же широте в Азии. Наоборот,ископаемые остатки из тропиков Азии указывают на то, что по крайней мере с третичного периода здесь климатические условия не изменились. Таким образом, мы должны отсюда сделать вывод, что изменение климата шло отнюдь не циркумполярно, но односторонне-асимметрично, так что Азия в отношении тропической зоны оставалась незатронутой этими климатическими изменениями. Вследствие этого, единственным возможным допущением для объяснения этого обстоятельства, надо признать перемещение климатических зон, вследствие изменения положения полюса.
На основании изложенного мы должны еще принять, для объяснения связи ареалов в том же растительном поясе, необходимость связанности материков. Так, если мы теперь бросим взгляд на южное полушарие, то и здесь увидим ряд разобщенных ареалов, охватывающих Ю. Америку и Австралию, а также Африку и Австралию или Ю. Америку, Африку и Австралию. Отсутствие захождения этих ареалов в северное полушарие не может быть случайным, и указывает на то, что эти ареалы являются частями когда-то сплошного распространения. Эта связь, напр. для Ю. Америки и Австралии, должна была осуществляться при посредстве самого короткого сообщения, т.-е. через Антарктику. Таким образом, мы должны принять связность материков южной умеренной зоны, которая должна была охватывать не только Ю. Америку, Австралию и Н. Зеландию, но также и Ю. Африку.
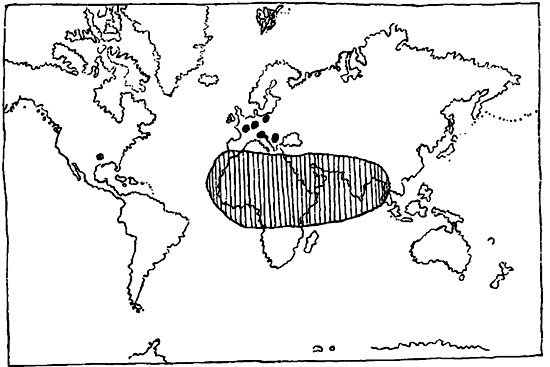
Теперь, на основании тех основных черт в характере ареалов покрытосемянных растений, которые мы только что установили, и тех выводов, которые мы должны были сделать, мы можем бросить взгляд на генезис этих ареалов. Из сказанного вытекает, что для многих семейств существовал такой период развития их ареалов, когда последние охватывали очень большие пространства, что требует допущения непосредственной связи Америки, Европы—Африки и Азии в пределах северной умеренной и тропической зон. Точно также заселение, в то время также связанной, умеренной зоны южного полушария требует возможности непосредственного переселения видов из Ю. Америки в Австралию и обмен формами между этой последней и Африкой. Это было время для многих семейств и родов наибольшей протяженности их ареалов.
Этот период образования ареалов мы можем считать 1-ой фазой их развития, для которой характерна необходимость иного расположения материков, чем теперешнее, и их большая связанность.
В последующее время эти ареалы подверглись изменениям под давлением различных влияний, из которых изменение климатических условий и расположения материков являются главнейшими. В результате этих изменений образовалась разорванность ареалов, их симметричность и зональность. Но в то же время роды, образующие эти ареалы, продолжали развиваться, возникали новые виды и роды, и распространение их создавало новые границы ареала. Вот эти вторичные моменты в развитии ареалов мы должны считать его 2-ой фазой. Для последней является характерным ясно выраженное приспособление ареалов к современному распределению материков и их отделенности один от другого. Здесь прежде всего характерно образование Атлантического океана, отделившего Европу от Америки; впервые появляется резкая разница между Старым и Новым Светом; на это же время падает окончательное отделение Ю. Америки и Австралии.
Вот теперь на основании изложенных данных мы можем перейти к вопросу о совпадении этих данных с теорией передвижения материков Вегенера, но прежде посмотрим, в каком отношении они будут находиться к гипотезе постоянства океанов. Сторонники последней, как уже было сказано, доказывают неизменность в распределении материков и океанов по крайней мере со времени появления сухопутной флоры и фауны. Как корректив к этой гипотезе явилась необходимость в допущении существования связывавших раньше современные материки участков суши, так называемых "мостов".
Изложенные выше данные, касающиеся 1-й фазы развития ареалов — ископаемые остатки тропических родов в Европе и Сев. Америке, доказанное переселение растений из Америки на Восток, закон зональности, — все это совершенно не может быть объяснено ни постоянством Атлантического океана, ни допущением северо-атлантического моста из Гренландии через Исландию в Шотландию. Здесь необходимо наличие материка, связывавшего Америку с Европой поперек Атлантического океана.
С другой стороны, 2-ая фаза развития ареалов вполне ясно очерчивает Атлантический океан, как преграду, возникшую в относительно позднее время, в промежутке между 1-й и 2-й фазой развития этих ареалов. Словом, при наличии постоянства океанов невозможно объяснить различную конфигурацию ареалов в их 1-й и 2-й фазе развития.
Другим обстоятельством, не поддающимся объяснению при постоянстве положения материков и полюса, требующих такого же расположения климатических зон как в настоящее время, является однородность некоторых ископаемых флор, требовавших для своего существования одинаковых климатических условий, которые в данное время однако оказываются расположенными в разных широтах. Между тем, если сблизить материки согласно теории Вегенера, то такие флоры окажутся расположенными в одних и тех же широтах.
Итак, теория Вегенера разрешает ряд вопросов генетической географии растений там, где гипотеза постоянства материков и океанов остается бессильной это сделать. Посмотрим дальше, насколько она согласуется с остальными из добытых Ирмшером данных. Мы видели, что многие формы в прежние геологические эпохи были распространены севернее, чем это позволяют современные климатические зоны, при чем это распространение отнюдь не было циркумполярно. Это обстоятельство может быть объяснено лишь асимметричным изменением климата, что может быть доказано лишь на основании рисуемого Вегенером перемещения полюсов.
Установленная выше симметричность ареалов, обусловленная пропуском в распространении в пределах меридиональной зоны Европа—Африка, может быть объяснена лишь на основе тех же перемещений полюсов, вызвавших именно в этой зоне резкие изменения климатических условий, повлекших за собой такие же резкие изменения флор. Начиная от мелового и кончая четвертичным периодом, климат Европы изменился от тропического до полярного с тем, чтобы после ледникового периода постепенно стать опять более теплым. Таким образом, симметричность ареалов в Америке и Азии представляет из себя прямой результат перемещения полюсов, результатом которого и является уничтожение большей части древней флоры в Европе, а вместе с этим и тех общих некогда черт во флоре Европы и Сев. Америки, и сохранение представителей этой флоры во всех широтах Азии и Америки.
Нам остается еще согласовать данные распространения покрытосемянных с основным положением Вегенера, касающимся перемещения самих материков. В этом отношении начерченная Вегенером картина положения материков в начале и в конце расхождения как нельзя более отвечает 1-й и 2-й фазам развития ареалов покрытосемянных. Своей гипотезой перемещения материков Вегенер не только делает понятной необходимость тесной связи С. Америки и Европы, но и объясняет до сих пор неразрешимую загадку распределения растительности южного полушария. Общность флоры южной Америки, Австралии, Антарктики и Африки, непонятная при теперешней их разобщенности, сразу разрешается возможностью обмена между их флорами при их прежней связанности в мезозое.
В заключение мы можем присоединиться к следующим выводам Ирмшера: 1) Для понимания географического распространения растений необходимо допущение перемещения полюсов и как его следствие — смещение климатических зон. 2) С данными распределения покрытосемянных лучше всего, из всех до сих пор высказанных точек зрения о происхождении материков, согласуется теория Вегенера, и, таким образом, эти данные могут служить доказательством и самой теории.
Нет никакого сомнения, что этим ботанико-географическим данным, как и самой гипотезе Вегенера, будут противопоставлены еще необъяснимые факты, и это уже имеет место главным образом со стороны геологов и геофизиков. Такие еще необъяснимые теорией Вегенера вопросы несомненно имеются и в биогеографии; так, мы можем указать, в качестве примера, хотя бы на еще почти незатронутую этой теорией проблему Тихого океана и происхождения флоры его островов. Но несомненно и то, что дальнейшее углубление и развитие мыслей Вегенера даст его гипотезе все новые и новые точки опоры. И нет ничего удивительного, что гипотеза Вегенера, гениальная по своей простоте, именно благодаря этой простоте, заставляющей невольно удивляться, как она до сих пор никому не пришла в голову, как это всегда бывало в таких случаях, создала себе ряд горячих сторонников и столько же страстных врагов. Но именно эта-то страстность и есть слабое место враждебной критики. И можно ли считать за таковую такого рода заявления, какие, например, делает известный палеоботаник Готан, говоря, что "учение Вегенера не есть уже геология или геофизика, а если можно так выразиться теософия, геометафизика или трансцендентная геология. Разница между приверженцами теории перемещения материков и полюсов и мною, — пишет Готан, — заключается в разнице воли к познанию и в различии установления границ возможности этого познания. Оба мы не знаем в действительности ничего о том, чем объясняются аномалии прежних времен; Вегенеровская школа лишь утверждает, что она это знает, тогда как я просто заявляю, что я (дальше известной грани) ничего не знаю".
Если бы человечество действительно остановило бы свои мысли на грани своего знания, то у нас не было бы науки. Но именно работа этой мысли, опережающей наши знания, и дает толчок к развитию последних, именно она способствует беспрестанному перемещению этой грани нашего познания, расширению нашего научного кругозора.
Эта опережающая добытые нами факты мысль и есть гипотеза. И поскольку гипотеза Вегенера базируется на научных фактах, нет никакого сомнения, что она открывает нам новые горизонты, создает широкие пути для развития биогеографической работы.
Ноябрь 1925 г.
1) Wegener A. Die Entstehung d. Kontinente u. Oceane, 2 And. (1922). Русский перевод Вегенер А. Происхождение материков и океанов (1925).
(стр. 53.)
2) Бельский П. Образование материков. "Природа" (1913). 991. Борисяк А. "Природа" (1919) и (1922).
(стр. 53.)
3) Svedelius. N. On the discontinuous geographical distribution of some tropical and subtrop. marine algae. Arch. f. Botan. 19 (1924); Koch, F. Ucber d. Rezcnte u. fossile Verbreit. d. Koniferen im Lichte neuerer geolog. Theorien. Mitt. d. d. dendrol. Ges. (1924), Nachträge lb. (1925); — Irmscher, E. Pflanzen Verbr. u. Entwicklung d. Kontinente. Mitt. d. Inst. f. allg. Botan. im Hamburg, V (1922). О последней см. реферат Б. А. Федченко. "Природа", № 1—6 (1924).
(стр. 59.)
4) Koch Fr. Die Cycadeen im Lichte der Wegenerschen Kontinent und Polwanderungstheorie. Mitteil. d. deutsch. Dendrolog. Ges. (1925).
(стр. 62.)