
"Природа", №07-09, 1919 год, стр. 327-350
Еще в начале восьмидесятых годов прошлого столетия, бывших временем расцвета чисто умозрительных теорий наследственности, было установлено понятие наследственной субстанции, т. е. такого вещества, которое, будучи скрыто в яйце или живчике, заключает в себе зачатки всех наследственных свойств будущего организма. В 1884 году О. Гертвиг и Страсбургер высказывают мысль, что таким носителем наследственных свойств организма является специально ядро половых клеток. Взгляд этот был принят затем известным теоретиком наследственности Вейсманном, по учению которого наследственная субстанция или, как он называл ее, зародышевая плазма состоит из ряда соподчиненных друг другу единиц: самые низшие из них или биофоры слагаются в детерминанты, из детерминантов образуются иды, а из ид — иданты. Последние, по его мнению, уже различимы в ядре с помощью микроскопа и идентичны с теми отдельностями, на которые распадается особое вещество ядра, его хроматин, во время процесса деления, (кариокинеза) и которые носят название хромозом. Как известно, число таких хромозом постоянно для каждого вида животных и растений, при чем иногда даже можно отличить по внешнему виду одну хромозому от другой, и вот по Вейсманну каждая такая отдельность заключает в себе совокупность различных единиц наследственной массы, при чем внутри этой высшей единицы наследственной субстанции или "иданта" следующие единицы или "иды" располагаются в линейном порядке (рис. 1).
Так сложилась теория наследственной монополии ядра, как ее многие называют, согласно которой ядро и в частности его хромозомы являются носителями наследственных свойств организма. Подобный взгляд скоро получил подтверждение с самых различных сторон: в пользу него говорили и явления, наблюдающиеся при делении клеток (кариокинезе), и процессы созревания половых продуктов и их оплодотворения, и, наконец, ряд опытных данных из области экспериментальной эмбриологии или механики развития, полученных, главным образом, Бовери, Гербстом и другими.
В течение девяностых и девятисотых годов против этой теории был сделан ряд возражений, стремившихся доказать, что не только ядро, но и другая главная часть клетки — протоплазма играет известную роль при передаче наследственных свойств. Этим однако отнюдь не было умалено значение хроматина, как носителя наследственной субстанции и в настоящее время, если и нельзя уже говорить о наследственной "монополии" ядра, то все же хромозомы следует признать главными носителями и передатчиками наследственных свойств, хотя бы некоторые особенности организмов и передавались при посредстве плазмы.
В начале настоящего столетия учению о хромозомах, как о главных носителях наследственных свойств, пришлось выдержать серьезное испытание, когда снова были открыты законы Менделя, при чем из этого испытания оно вышло с честью, доказав тем свою жизнеспособность.
Как известно, основной сущностью законов Менделя является положение, что при скрещивании различных в каком-либо отношении форм особенности родителей не сливаются друг с другом в половых клетках помеси, а последняя образует два сорта половых клеток — одни с особенностью матери, другие с особенностью отца, отчего в дальнейшем и наступает столь характерное для менделистической наследственности расщепление. Если хромозомы половых клеток, действительно, заключают в себе наследственные свойства будущего организма, то должна быть определенная связь между этим менделистическим расщеплением, наблюдаемым в опытах скрещивания, и взаимоотношениями хромозом во время созревания половых продуктов и оплодотворения. И вот уже в начале девятисотых годов благодаря работам Монтгомери, Сеттона и Бовери удалось создать гипотезу, вполне удовлетворительно объясняющую все явления расщепления, при помощи известных перегруппировок хромозом. (См. статью С. Е. Кушакевича в "Природе" 1913 г. № 10, стр. 1205—1220).
Вскоре вслед за этим, главным образом, американскими исследователями (Мэк Кленг, Вильсон, Стивенс и др.) были открыты и изучены особые хромозомы, получившие название гетерохромозом или половых хромозом, так как они имеют самую тесную связь с определением пола 1), наследование которого совершается также по законам Менделя и обнаруживает снова наглядную связь между данными цитологии и менделистического наследования. — Однако при своих исследованиях цитологи имели дело с одними объектами, а экспериментаторы-менделисты совершенно с другими, так что не удавалось проверить все эти данные на одном и том же животном или растении. "Мы должны, писал в 1909 году Гэккер, найти прежде всего такие объекты, которые будут одинаково удобны как для целей цитологического исследования, так и для работ по физиологии наследственности. В настоящее же время между коньюгацией гетерохромозом у клопов и явлениями расщепления у горохов и мышей еще лежит обширная область неясностей и произвольных толкований".
В настоящее время такой объект найден в лице американской плодовой мухи Drosophila ampelophila, которая неустанно изучается в Америке с 1910 года, главным образом, Морганом и его ближайшими сотрудниками, при чем опубликован о наследственности у этой формы ряд интереснейших работ. В 1915 году этими исследователями была выпущена книга "Механизм менделистической наследственности", представляющая собою сводку полученных ими результатов, с данными которой, а также некоторых других работ этих авторов мы и познакомим здесь читателя 2).
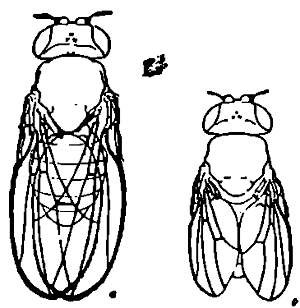
Скажем однако прежде всего два слова об объекте всех этих исследований. Мелкая плодовая муха Drosophila ampelophila из семейства Drosophilidae относится к наименее изученной группе мух "Асаlyptrata". Главный интерес этой формы заключается в том, что, начиная с 1910 г., она производит в лаборатории Моргана ряд новых форм путем внезапных изменений или мутаций. Эти новые формы или мутанты отличаются от произведшей их основной или "дикой" формы самыми различными особенностями: цветом тела, цветом глаз, размерами и строением крыльев и т. п. На нашем рис. 2 изображен рядом с нормальной Drosophila ampelophila ("дикая форма" по выражению Моргана) ее мутант с миниатюрными крыльями. Благодаря появлению таких новых форм явилась возможность изучать возникавшие при этом признаки путем скрещивания, так что в настоящее время изучена наследственность не менее чем ста признаков как нормальной формы, так и ее мутантов.

Чрезвычайно интересна Drosophila ampelophila и в цитологическом отношении. Как показали исследования главным образом Метца у этой формы и у некоторых близких к ней видов имеется 4 пары хромозом (см. рис. 3): одна пара половых хромозом, несколько различных у самца и самки (h), две пары обыкновенных хромозом или "аутозом" (l) и одна пара мелких микрохромозом (m). Половые хромозомы у различных форм бывают двоякого рода: в одних случаях самка имеет пару таких образований, а самец одно — тогда говорят о гетерохромозомах или Х-хромозомах, в других же случаях оба пола имеют по 2 половых хромозомы, но у самца один из этих элементов недеятелен и находится как бы в состоянии, близком к рудиментарному — в этом случае говорят об идио-хромозомах и обозначают деятельную из них, как X, а недеятельную, как Y, так что самка при этом является XX, самец же — XY. Как раз последний случай мы и имеем у Drosophila, как это хорошо видно на рис. 4, где Y-хромозома у самца чисто условно изображена крючковидной формы. Мы стоим, таким образом, перед вопросом, нельзя ли связать ряд внешних особенностей Drosophila, наследование которых уже изучено, с этими цитологическими отношениями, что, действительно, и удалось Моргану и его школе.

Дело облегчается здесь тем обстоятельством, что часть особенностей Drosophila ampelopbila и ее мутантов наследуется в тесной связи с полом — по типу так называемой ограниченной полом наследственности. Именно эти случаи позволили Моргану и Гольдшмидту еще в 1911 году установить их теорию определения пола, согласно которой имеются особые зачатки или факторы для мужского и женского пола, при чем один из них локализован в половых хромозомах — именно в Х-хромозоме. Согласно той же теории Моргана в Х-хромозомах заключены и зачатки или факторы всех тех особенностей, которые наследуются в связи с полом, что и объясняет своеобразный ход их наследования.

Возьмем одну из последних — именно передачу по наследству белого цвета глаз у особой мутации Drosophila, которому противоположен (составляет с ним пару или аллеломорфу на языке менделистических символов) красный цвет глаз обыкновенной мухи. При скрещивании последней с белоглазой формой наследование идет различно в зависимости от того, имелись ли белые глаза у самца или у самки. В первом случае (рис. 5) красный цвет глаз доминирует в первом поколении помесей, во втором же происходит обычное расщепление на красных и белых в отношении 3:1. Во втором случае, т. е. когда белоглазой формой является самка, в первом поколении все самцы имеют белые глаза, а все самки — красные, во втором же поколении среди представителей обоих полов наблюдается поровну белоглазых к красноглазых форм (рис. 6).
Для объяснения всех этих отношений достаточно принять, что зачатки данных особенностей заключены в Х-хромозомах, которых у самки имеется 2, а у самца только 1, так как другая половая хромозома или Y-хромозома, свойственная представителям мужского пола, не заключает в себе никаких зачатков. На рис. 5 и 6 изображен ход наследования и этих хромозом, при чем заключающие фактор красного цвета глаз обозначены черным, а содержащие фактор противоположной особенности белым, и из рассмотрения этих рисунков весь ход наследования в обоих случаях становится совершенно ясным, если принять, что в случае соединения двух Х-хромозом получается самка, а при соединении X и Y-хромозомы самец.
Красный и белый цвет глаз не являются единственными особенностями, наследуемыми у Drosophila в связи с полом: число последних очень велико — достигает до 50. Сюда относятся некоторые другие цвета глаз (например, вишневый, эозиновый), желтая окраска тела, рудиментарное и миниатюрное развитие крыльев и многие другие. Зачатки, факторы или гены (как их часто теперь называют) этих особенностей следует признать благодаря этому локализованными в той паре хромозом, которые были названы выше половыми хромозомами (рис. 3 h, рис. 4 XX и XY).
К чему же должна приводить локализация зачатков известных особенностей в одной и той же хромозоме или паре хромозом? Конечно, такие особенности должны передаваться потомству в самой тесной связи друг с другом, как будто бы дело шло при этом не о различных свойствах организма, а об одном, но лишь выражающемся в различных чертах строения одной и той же особи. Подобную связь различных особенностей друг с другом обыкновенно называют корреляцией или коррелятивной зависимостью, если же дело идет о связи некоторых признаков при наследовании, когда они бывают первоначально скрыты в половых клетках или гаметах, то это явление называют гаметической корреляцией. Таким образом, все наследуемые по типу ограниченной полом наследственности особенности Drosophila ampelophila благодаря локализации их генов в половых хромозомах обнаруживают гаметическую корреляцию или, как иногда говорят, гаметическое сцепление друг с другом.
Заметим, что эти явления у нашей мушки не представляют собою чего-либо существенно нового, так как гаметическая корреляция была открыта еще в 1906 году Бэтсоном и Пеннеттом у душистого горошка, а затем такие же случаи стали известны и у других растений и животных. Однако самая сущность этого явления была совершенно непонятна, и только Моргану удалось выяснить, что явления гаметической корреляции свидетельствуют о локализации генов в одних и тех же хромозомах.
Однако у Drosophila, как мы видели выше, кроме половых имеется еще 3 пары хромозом: 2 пары обыкновенных или "аутозом" и 1 пара рудиментарных "микро-хромозом"; нельзя ли обнаружить локализацию каких-либо особенностей и в этих хромозомах?
Первоначально Морган и его сотрудники имели дело с корреляцией или "сцеплением" только тех особенностей, которые наследуются в связи с полом, однако несколько позже такое же "сцепление" удалось найти и между теми особенностями, наследование которых связи с полом не имеет, так что их гены, очевидно, помещаются не в половых хромозомах.
Благодаря работам Моргана и Линча (1912), Стёртеванта (1913), Стёртеванта и Бриджа (1914) здесь удалось обнаружить уже не одну, а две самостоятельных группы особенностей, при чем члены каждой такой группы обнаруживают корреляцию или "сцепление" лишь друг с другом, но не с членами других групп. Отсюда уже вполне естественно было сделать вывод, что гены этих особенностей локализованы в других парах хромозом — скорее всего в первой и во второй паре аутозом или обыкновенных хромозом, изображенных также на рис. 3.(l) и 4. — Число особенностей, присущих каждой такой группе, также довольно велико и превышает теперь уже два десятка; к одной из них относятся такие признаки, как черный цвет тела, пурпуровые глаза, недоразвитые крылья и др., к другой — тоже своеобразный черный цвет тела (эбеновый), розовые глаза, четковидные крылья и т. д.
Наконец, в 1914 году Муллеру и Ходжу удалось найти два признака, гены которых по тем же причинам следует признать находящимися в четвертой паре хромозом, именно в маленьких микрохромозомах (рис. 3 m и рис. 4) — это, во-первых, изогнутость крыльев и, во-вторых, безглазость. Наполовину рудиментарный характер микрохромозом не позволяет думать, чтобы число членов этой группы могло бы сильно возрасти при новых исследованиях.
Таким образом, эти исследования Моргана и его школы приводят нас к установлению полной аналогии между группировкой наследственных свойств и строением хромозом, доказывая лишний раз, что последние, действительно, можно признать носителями наследственных свойств. В то же время эти данные разрешают окончательно разногласие между Вейсманном, полагавшим, что в каждой хромозоме заключены все наследственные свойства данной особи, и Бовери, выдвинувшим гипотезу качественного различия хромозом, согласно которой каждая из них отличается не только по своему внешнему виду, но и по своему "наследственному составу", т. е. что одна хромозома является носителем одних, а другая других свойств. Изложенные здесь данные о локализации генов у Drosophila, конечно, решительно говорят в пользу последнего воззрения.
Еще интереснее, что на основании тех же самых исследований над наследственностью различных особенностей у Drosophila можно составить себе известное представление и о расположении генов в каждой такой хромозоме. Путь к этому открывают явления так называемого "перекреста" (crossing over) и обмена частицами между различными хромозомами, изученного отчасти Морганом, а, главным образом, Стёртевантом (1913, 1915). Явление это уже довольно сложно, почему мы разберем его здесь лишь в самых общих чертах.
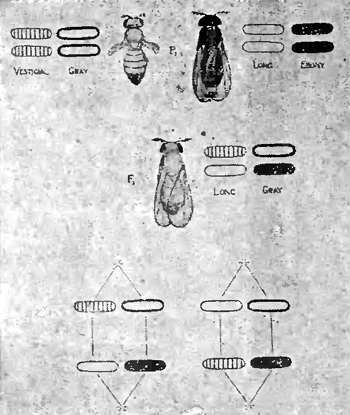
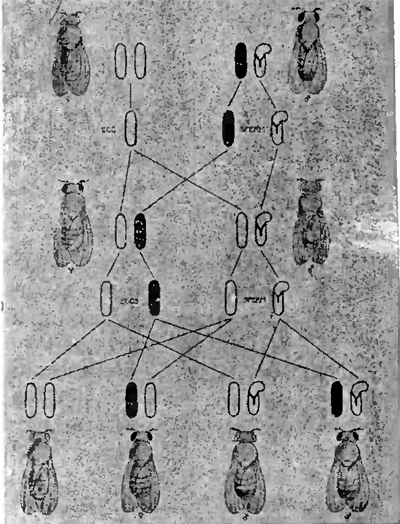
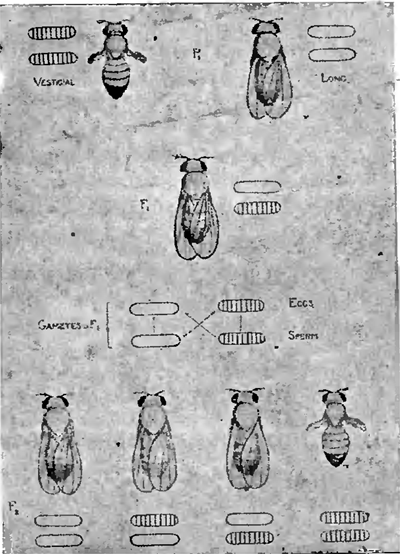
Как известно, между явлениями менделистического расщепления и перегруппировками хромозом во время процессов созревания и оплодотворения яйца имеется несомненная связь. Последняя наглядно выступает на наших рис. 7 и 8, изображающих ход наследования по Моргану одной или двух пар особенностей, которыми только и отличались друг от друга исходные формы. В первом случае (рис. 7) это отражается только на одной какой-нибудь паре хромозом, почему в результате и возникает более простое (моногибридное) расщепление на две группы особей в отношении 3:1. Во втором случае (рис. 8) замешаны уже две пары хромозом, почему и расщепление принимает более сложный (дигибридный) характер, образуется 4 сорта половых клеток и возникает четыре группы различных форм в отношении 9:3:3:1, что изображено в виде диаграммы на рис. 9.
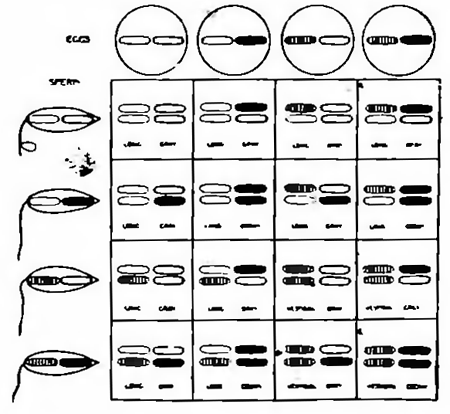
Так обстоит дело, если скрещиваются особи, отличающиеся двумя парами признаков, как это имело место в последнем примере (недоразвитые крылья — длинные крылья, серый цвет — черный цвет), причем каждая такая пара противоположных признаков локализована в особой паре хромозом. Что будет однако, если мы скрестим формы, отличающиеся друг от друга также двумя парами признаков, но уже такими, гены которых сосредоточены в одной и той же паре хромозом (будь то в половых или в одной из пар аутозом)?
Конечно, здесь должны иметь место уже иные отношения, так как выше уже говорилось, что о локализации известных генов в одной и той же паре хромозом и судят по тому, что вызываемые ими внешние особенности обнаруживают при передаче по наследству известное "сцепление" или корреляцию.
Если бы гаметическое сцепление или корреляция таких генов была бы полной, то в этом случае две пары данных свойств наследовались бы, как одна, по типу моногибридного скрещивания (см. рис. 7). Однако совершенно полной гаметической корреляции или "сцепления" обыкновенно все же не бывает, и дело протекает здесь уже не так просто.
Предположим, у нас происходит скрещивание двух Drosophila, у одной из которых крылья недоразвиты (как на рис. 7 и 8) и цвет тела черный, но не цвета эбенового дерева, как на рис. 8 (особенность, ген которой локализован во второй паре аутозом), а чисто черный (его ген находится вместе с геном недоразвитых крыльев в первой паре аутозом), у другой же мухи крылья нормальные и цвет тела серый. В первом поколении проявляются (доминируют) последние особенности (нормальные крылья и серый цвет), во втором происходит менделистическое расщепление. Если бы оно было чисто моногибридным (а "сцепление" совершенно полным), то возникло бы лишь две группы форм, подобных исходным в отношении 3:1 (как на рис. 7). Если бы "сцепления" совсем не было, т. е. гены занимали бы места в различных хромозомах, то получилось бы типичное дигибридное расщепление на 4 группы форм (две, как исходные, и две новых) в отношении 9:3:3:1 (как на рис. 8 и 9). На самом деле получается нечто промежуточное между тем и другим случаем: громадное большинство форм второго поколения (как это вообще характерно для случаев гаметической корреляции) представляет возврат к типу исходных форм, но кроме них имеются еще две новых группы форм (недоразвитые крылья и серый цвет, нормальные крылья и черный цвет), представленные однако значительно меньшим числом особей, чем это бывает при нормальном дигибридном расщеплении. Очевидно, здесь имеется "сцепление" между генами, но не настолько полное, чтобы возникало только два сорта половых клеток, как при моногибридном расщеплении (рис. 7), а получается их все же четыре сорта, как при дигибридном (рис. 8). Однако в последнем случае, когда никакого сцепления нет и гены локализованы в различных хромозомах, все четыре сорта гамет образуются в равном количестве (см. рис. 8 и 9), здесь же, как это обычно бывает при гаметической корреляции, двух сортов гамет получается гораздо больше, чем других, и в частности гамет с генами нормальных крыльев и серого цвета и недоразвитых крыльев и черного цвета (т. е. с особенностями исходных форм) получается гораздо больше, чем промежуточных между ними.
Есть даже способ, чтобы определить, в каком отношении у гибридов первого поколения образуются различные свойственные им сорта гамет. Для этого нужно скрестить такого гибрида с чистой рецессивной формой в смысле всех ее особенностей, так как такие формы, как известно, производят только один сорт гамет. В данном случае такой формой будут черные мухи с недоразвитыми крыльями (оба этих признака исчезают в первом поколении — значит, они рецессивны). Производя подобное скрещивание получаем среди самцов две группы форм (подобные исходным), среди самок четыре группы форм (две подобные исходным, две промежуточных между ними). Отсюда приходится сделать вывод, что у самцов имеется только 2 сорта гамет, у самок же 4; об отношении же последних можно судить по отношению получающихся при этом особей. Это отношение таково:
| Черные недо- развитые |
Серые нор- мальные |
Черные нор- мальные |
Серые недо- развитые |
| 41,1 | 41,5 | 8,5 | 8,5 |
т. е. каждого сорта особей, подобных исходным, приблизительно в 5 раз больше, чем промежуточных между ними. Очевидно и гамет у самки первого поколения гибридов образуется 4 сорта в отношении 41,5 : 41,5 : 8,5 : 8,5.
То же самое явление наблюдается во всех случаях, когда дело идет о скрещивании особей Drosophila, отличающихся двумя парами признаков, но такими, которые обнаруживают гаметическое сцепление, почему предполагается, что их гены локализированы в одинаковых хромозомах. Как показывает указанный сейчас способ, при этом обыкновенно оказывается, что самцы образуют всегда 2 сорта гамет, как будто бы данное скрещивание было моногибридным, самки же 4 сорта, как при дигибридном скрещивании, но не в равном друг другу отношении, а всегда с значительным перевесом тех гамет, которые образуются самцом. Числовые отношения получаются при этом различные и изменяются в зависимости от того, с какими особенностями мы имеем дело. В приведенном выше примере отношение нормальных гамет к промежуточным ("неперекрещенных" к "перекрещенным", по выражению Моргана) было 41,5:8,5 или лучше, если привести эти цифры к 100 — 83:17. Вот еще некоторые другие цифры из опытов Моргана и его сотрудников:
| Отношение гамет: | |||
| Хромо- зомы. |
Особенности. | нормальных или "непере- крещенных" |
промежу- точных или "перекре- щенных" |
| Половые: | желтый цвет | 99 | 1 |
| белые глаза | |||
| " | белые глаза | 67 | 33 |
| миниатюрн. крылья | |||
| " | желтый цвет | 57 | 43 |
| рудимент. крылья | |||
| 1 п. ауто- зом. |
черный цвет | 83 | 17 |
| недоразв. крылья | |||
| " | недоразвитые и | 92 | 8 |
| изогнутые крылья | |||
| 2 п. ауто- зом. |
цвета сепии | 75 | 25 |
| розовые глаза | |||
| " | розовые глаза | 52 | 48 |
| цвета эбенов. дер. | |||
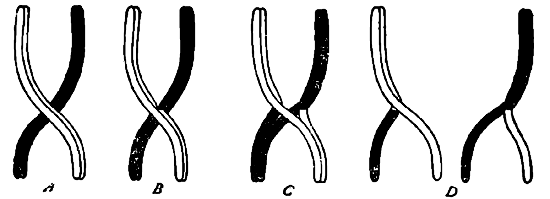
Чем же объяснить, как самое появление этих гамет с промежуточными особенностями, так и возникающие при этом различные числовые отношения? Для объяснения этого явления Морган еще в 1911 году предложил гипотезу, согласно которой между гомологичными хромозомами происходит обмен частицами, в результате которого кроме двух первоначальных сортов хромозом получаются еще два новых, состоящих наполовину из частиц одной хромозомы и наполовину из частиц другой, т. е. имеющих промежуточный характер. Самый механизм этого процесса он представляет себе таким образом, что на известной стадии развития половых клеток гомологичные друг другу хромозомы, полученные в свое время одни от матери, другие от отца, соединяются и переплетаются друг с другом, а затем расходятся, при чем одна уносит уже часть другой, как это изображено наглядно на рис. 10. Этот процесс Морган и назвал перекрестом (crossing over), считая его основной причиной описанной выше гаметической корреляции.

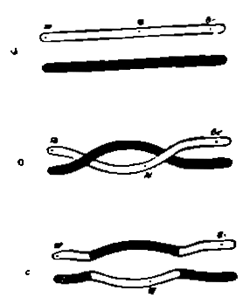
Нельзя не отметить, что он руководился при создании данной гипотезы чисто фактическими отношениями, описанными незадолго до того Янссенсом при сперматогенезе у земноводного Batracoseps attennatus и легшими в основу развитой этим автором теории хиазматипии 3). Янссенс, как и другие иследователи, наблюдал во время особого периода развития семенных клеток (в период роста и превращения сперматогоний в сперматоциты) соединение и переплетание друг с другом тех тонких нитей, в которые соединяются при этом хромозомы этих клеток. Для объяснения всех наблюдающихся при этом картин он и предполагает, что здесь происходит соединение или коньюгация друг с другом гомологичных хромозом, которые соединяются крест на крест в виде греческой буквы χ. (хи), а затем расходятся, расщепляясь кроме того во время этого процесса вдоль (рис. 11). Как видно на последнем рисунке, при этом получается в конце концов 4 хроматиновых элемента (D), распределяющиеся по особым гаметам, при чем в двух таких элементах (и, тем самым, гаметах), обмена частицами не произошло ("непекрещенные" гаметы), в двух же других он имел место ("перекрещенные" гаметы). Это и объясняет вполне существование отмеченных выше четырех сортов половых клеток.
Чем же можно объяснить, наконец, их изменчивое отношение друг к другу в различных случаях? Еще в 1911 году Морган высказал предположение, что это зависит от большей или меньшей близости генов в хромозомах друг к другу, более же детально эта идея была разработана Стёртевантом (1913, 1915). Согласно последнему все гены занимают в хромозомах строго определенное положение, будучи расположены в линейном порядке, при чем отношение "перекрещенных" к "неперекрещенным" гаметам или процент перекреста зависит от расстояния между двумя данными факторами в хромозоме.
Эта идея не может считаться чистой гипотезой, так как ее до известной степени можно проверить и опытным путем, именно, зная расстояние между факторами А и В и между В и С, можно попытаться предсказать также и расстояние между первым и последним из них, т. е. между А и С. В самом деле, если все эти гены расположены по одной линии, то третий из них С может лежать или между А и В или дальше за В. В первом случае, чтобы определить расстояние между А и С (или АС), нужно из расстояния между А и В (или АВ) вычесть расстояние между В и С (или ВС), во втором же случае эти величины нужно сложить друг с другом, т. е. АС = АВ ± ВС.
Применим теперь этот расчет к приведенным выше цифрам. Процент перекреста и, значит, расстояние между генами желтого цвета и геном белых глаз в половых хромозомах (АВ) = 1, эта же величина между геном белых глаз и геном миниатюрных крыльев (ВС) = 33, отсюда заключаем, что расстояние между последним геном и геном желтого цвета (АС) = 33 ± 1 и, действительно, оно близко к 34, как показывает специальный опыт на основании получающегося при этом процента перекреста.
Другой пример: выше же были даны эти величины для факторов черного цвета и недоразвитых крыльев (17) и для факторов недоразвитых и изогнутых крыльев (8), локализованных в первой паре аутозом; отсюда без труда заключаем, что процент перекреста и расстояние между факторами черного цвета и изогнутых крыльев может равняться 17 ± 8, и, действительно, оно равно 25.
Однако дело обстоит не всегда так просто и гладко и иногда между ожидаемыми и наблюдаемыми цифрами замечается резкое различие. Так, например, выше была указана для процента перекреста и расстояния между факторами желтого цвета и рудиментарных крыльев цифра 43 — теоретически же следовало ожидать значительно большую (именно 55), наоборот, для генов розовых глаз и цвета эбенового дерева должна бы быть меньшая цифра — вместо 48 только 30 и т.д. Стёртевант находит выход из данного затруднительного положения в допущении, что в подобных случаях на самом деле замешано не две пары, а три пары факторов, между которыми и происходит двойной перекрест. Сущность последнего явления изображена наглядно на рис. 12, детали же его отличаются уже значительной сложностью, что и дало повод Троу 4) попытаться этим путем подорвать значение всей теории. Безусловно, всякие вспомогательные гипотезы всегда несколько вредят цельности каждой теории, но, с другой стороны, нельзя никогда и требовать от природы, чтобы она была проще, чем есть на самом деле. Вот почему мы не видим причин отвергать эту дополнительную гипотезу Стёртеванта, объясняющую многие сложные случаи.
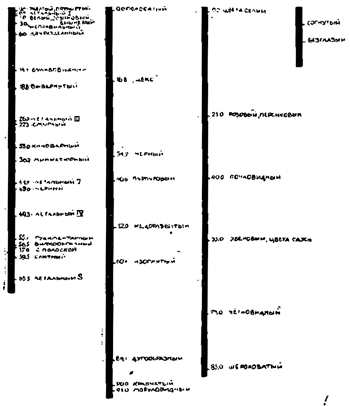
Как бы то ни было, на основании всех этих соображений и целого ряда опытов Морган и его сотрудники не ограничиваются данными о локализации тех или иных факторов в известных хромозомах и констатированием расположения этих генов внутри хромозом в линейном порядке, а дают в виде конечного вывода всей их работы диаграмму, наглядно изображающую эту локализацию по крайней мере для наилучше исследованных генов. Она воспроизведена на рис. 13, при чем первой слева помещена половая (X-) хромозома и в ней показаны места расположения 22 факторов, за ней следует первая и вторая аутозомы, в которых также намечены места 17 факторов, и, наконец, справа мы видим микрохромозому с ее двумя факторами. Цифры показывают расстояние (в %) каждого гена от точки, принятой за начальную.
Нельзя не согласиться со словами Стёртеванта, которыми он заканчивает свою первую работу по данному вопросу: "все эти факты, говорит он, являясь новым аргументом в пользу хромозомальной теории наследственности, доказывают — теперь уже чисто математически — что наследственные факторы расположены в хромозомах в линейном порядке".
На основании изложенных данных приходится во многом изменить наше представление о сущности менделистических факторов и о способе возникновения в процессе эволюции новых свойств, на чем мы и должны сейчас остановиться.
Одной из важнейших вспомогательных гипотез современного менделизма была до последнего времени так называемая гипотеза присутствия-отсутствия, развитая Бэтсоном еще в его первых работах (1902, 1906). Согласно последней наличность в половых клетках особого детерминанта, т. е. определителя, как выражался в свое время Вейсманн, или же фактора, иначе гена, как мы говорим теперь, обязательна отнюдь не для всех наследственных свойств организма, а только для одной части их — именно для тех, которые проявляются или доминируют в первом поколении помесей, те же особенности, которые у гибридов первого поколения исчезают, иначе говоря рецессивны, возникают по Бэтсону при отсутствии в гаметах соответствующих зачатков или генов. Новые особенности или новые свойства, согласно господствующим в настоящее время воззрениям, возникают в процессе эволюции путем внезапных изменений или мутаций, открытых в 1900 году Коржинским и де-Фризом. В согласии с теорией присутствия-отсутствия и мутации до последнего времени разделялись на две главных группы: прогрессивные, когда к старым прибавляется новое свойство и новый ген, и регрессивные, когда одно из старых свойств и его фактор исчезают.
Однако в самое последнее время против как теории присутствия-отсутствия, так и деления мутаций на прогрессивные и регрессивные стали раздаваться отдельные голоса 5), и окончательный удар им был нанесен Морганом в его теории тождественных мест, возникшей в результате изложенных выше исследований над Drosophila.
При скрещивании различных рас Drosophila Моргану пришлось столкнуться с некоторыми фактами, которые совершенно не гармонируют с теорией присутствия-отсутствия. Отметим здесь лишь один из них. При скрещивании красноглазой самки Drosophila с белоглазым самцом, как отмечалось уже выше, в первом поколении красный цвет доминирует, а во втором происходит расщепление на 3 красноглазых и 1 белоглазую особь (см. выше рис. 5). Если самка Drosophila, имеющая глаза цвета эозина скрещивается с белоглазым же самцом, наблюдается подобное же доминирование цвета эозина над белым с последующим расщеплением в отношении 3:1. Теория присутствия-отсутствия может объяснить это тем, что красный глаз зависит от одного фактора (R), цвет эозина от другого фактора (Е) и белый цвет возникает при отсутствии этих двух факторов, почему и красный цвет и цвет эозина доминируют над белым. Следовательно, на языке букв красноглазая Drosophila будет Re (есть фактор красного цвета, нет фактора цвета эозина), муха с эозиновыми глазами — rE (наоборот), белоглазая Drosophila — re (нет обоих факторов). Если это, действительно, так, то при скрещивании красноглазой формы с имеющей глаза цвета эозина различие между исходными формами должно касаться 2 факторов (Re и rE) и быть дигибридным с расщеплением во втором поколении на 4 группы форм в отношении 9:3:3:1. Однако в действительности ничего подобного не наблюдается и оба этих цвета ведут себя тоже так, как будто бы они составляли одну пару, т. е. в первом поколении помесей красный цвет доминирует над эозиновым, а во втором происходит расщепление на 3 красных и 1 цвета эозина. Словом, с точки зрения теории присутствия-отсутствия три данных свойства должны были бы составить 2 пары или 2 двойных аллеломорфы (красный—белый, эозиновый—белый), на самом же деле этого нет и получается система тройной аллеломорфы (красный—эозиновый—белый), где каждый предыдущий доминирует над всеми последующими, что совсем не предусматривается теорией присутствия-отсутствия и даже резко противоречит ей.
Морган приводит такие же примеры множественных аллеломорф, как он их называет (при чем они могут быть не только тройными, но четверными и даже более) и у других животных и растений (например, у кроликов,шелковичного червя, бобов, Lychnis и др.) и дает всем этим случаям очень простое объяснение при помощи названной выше теории идентичных мест.
Согласно Моргану теория присутствия-отсутствия, допускающая лишь две возможности — присутствие и отсутствие данного фактора, глубоко ошибочна. "Предполагать об отсутствии фактора на основании отсутствия внешней особенности, говорит он, столь же наивно, как думать, что животное двигается к свету потому, что любит последний". Действительно, раз чисто материально каждый ген представлен лишь известной точкой в той или иной хромозоме, то не проще ли допустить, что изменение любого пункта хромозомы, т. е. заключающегося там гена, возможно во многих, при том самых различных направлениях, а не только в двух, условно обозначаемых как положительное (присутствие) и отрицательное (отсутствие).
Если мы обратимся теперь к схеме расположения генов Drosophila в ее хромозомах (рис. 13), то увидим, что в некоторых точках последних помещается по 2 и даже по 3 гена. Так, в пункте 1,0 половой хромозомы локализованы гены белого, эозинового и вишневого цвета глаз, во второй аутозоме в точке 25,0 заключены гены розового и персикового цвета глаз, а в точке 55,0 ее находятся гены двух темных окрасок всего тела — эбенового и цвета сажи. Всем этим особенностям, как и всем изображенным на данной схеме, противоположны соответствующие особенности нормальной мухи (или "дикой формы" по выражению Моргана) — красный цвет глаз, серая окраска тела и пр. Очевидно, фактор или вернее факторы последней занимали те же места в хромозомах, которые заняты теперь факторами этих произошедших от нормальной мухи мутационным путем новых форм или рас Drosophila. Значит, мутационная изменчивость последней и вызывалась прежде всего тем, что соответствующие или идентичные места той или иной хромозомы (откуда и термин — теория тождественных или идентичных мест) подверглись каким то тончайшим изменениям структурного или химического характера. Наличность же у различных форм в одном и том же пункте той или иной хромозомы (как в 1,0 у половых хромозом, в 25,0 и в 55,0 во второй паре аутозом) нескольких генов наглядно показывает, что во время мутационного процесса изменения совершались в нескольких различных направлениях. Таким образом, принадлежность двух или нескольких признаков к одной аллеломорфе (обыкновенной двойной или более редкой множественной) объясняется не присутствием или отсутствием гена, а нахождением соответствующих генов в идентичной точке хромозомы, что говорит и за их происхождение друг от друга мутационным путем. Словом, теория идентичных мест Моргана совершенно вытесняет теорию присутствия-отсутствия Бэтсона, и тем самым отпадает деление мутаций на прогрессивные и регрессивные, как совершенно ненужное и создающее только безполезные ограничения для нашего понимания эволюционного процесса.
Таким образом, согласно той же теории, основной причиной различных наследственных изменений организмов или мутаций, как их теперь называют, на которые в конечном итоге сводится вся эволюция живых существ, следует признать какие то тончайшие изменения в хромозомах.
К последнему выводу можно притти и чисто дедуктивным путем: раз хромозомы, действительно, являются носителями наследственных свойств, то каждое наследственное изменение или мутация должна непременно сопровождаться тем или иным изменением в хромозомах. Однако все дедукции этого рода гораздо менее доказательны, чем одно точно установленное наблюдение, как то, на котором и основывается сделанный выше вывод.
Этот вывод подтверждается кроме того, и чисто цитологическим исследованием представителей той растительной формы, у которой впервые было открыто и изучено явление мутаций, т. е. у Oenothera lamarckiana.
За последнее время ряду исследователей удалось установить, что и здесь мутационная изменчивость сопровождается некоторыми изменениями в хромозомах — именно в их числе 6).
Согласно наблюдениям целого ряда исследователей не только у энотеры Ламарка, но и у многих других видов того же рода число хромозом в клетках тела (так называемое двойное или диплоидное число) равно 14 — в половых же клетках (так называемое простое или гаплоидное число) — 7. Однако в 1907 году мисс Лютц удалось обнаружить, что один из мутантов энотеры, именно гигантская энотера (Оеп. gigas), имеет вдвое большее число хромозом, т. е. содержит в гаметах 14 и в клетках тела 28. Подобные случаи удвоения числа хромозом известны и у других форм как из животного, так и из растительного царства, при чем подобные формы получили название тетраплоидных.
Очевидно, нормальные (диплоидные) энотеры производят гаметы с 7 хромозомами, гигантская же энотера — с 14. Не трудно предвидеть, что при скрещивании Oen. gigas с любой другой энотерой возникнет помесь, имеющая уже не 14 и не 28, а 21 хромозому, и, действительно, в 1912 году подобные триплоидные растения, как их можно назвать, были получены также мисс Лютц и Стомпсом.
Еще более интересно открытие, сделанное в том же году мисс Лютц и Гэтсом, что у особого мутанта энотеры — Oen. lata — имеется всегда не 14, а 15 хромозом, т. е. здесь удвоению подвергся не весь состав хромозом, как у гигантской энотеры, а лишь один из его членов.
Относительно толкования этих наблюдений мнения несколько расходятся. Стомпс 7) и с ним некоторые другие думают, что измененное число хромозом есть само только следствие мутации, напротив, по мнению Гэтса и других, эти изменения в числе хромозом являются непосредственной причиной мутаций. Как нам кажется, последний взгляд более обоснован и имеет гораздо более за себя, и вообще мысль о самой тесной и при том причинной связи между мутационной изменчивостью, с одной стороны, и между известными изменениями в числе или тончайшей структуры хромозом, с другой, нельзя не признать в высокой степени вероятной.
Таким образом, взгляд, высказанный еще в 1884 году О. Гертвигом и Страсбургером, за протекшие с тех пор 35 лет получил целый ряд чрезвычайно существенных подтверждений. Изложенные наблюдения школы Моргана над Drosophila и некоторых других исследователей над Oenothera особенно важны потому, что они перебрасывают мост между теми двумя областями (чисто цитологический и экспериментально-генетической), которые развивались до сих пор независимо друг от друга. Совпадение же двух независимых друг от друга свидетельских показаний, как известно, имеет всегда особенное значение для установления действительного существования какого-либо факта, почему теперь мы можем считать его уже гораздо более доказанным, чем раньше.

1) См. статью В. М. Шимкевича об определении пола у животных в "Природе" за 1915 год (№№ 1 и 2). (стр. 330.)
2) Morgan, Sturtevant, Muller, Bridges. The mechanism of mendelian heredity. New Jork. 1915.
Важны также статьи тех же авторов: Morgan в Journ. Exp. Zool. 11 (1911), Morgan and Lynch в Biol. Bull. 23 (1912), Sturtevant в Journ. Exp. Zool. 14 (1913), Sturtevant в Science 37 (1913), Bridges and Sturtevant в Biol. Bull. 26 (1914), Sturtevant в Zeit. ind. Abst. Ver. 13 (1915), Muller в Journ. Exp. Zool. 17 (1914), Hoge в Amer. Natur. 49 (1915), Muller в Amer. Nat. 50 (1916), а также специально о хромозомах у Drosophila статьи Metz в Journ. Exp. Zool. 17 (1914) и 21 (1916). (стр. 331.)
3) Janssens. La theorie de la chiasmatypie. — Cellule 25. 1909. (стр. 342.)
4) Trow. A criticism of the hypothesis of linkage and crossing over. Journ. Getretics. V. № 4. 1916. (стр. 344.)
5) См., например, веские замечания Баура во втором издании его книги о наследственности. (стр. 346.)
6) Хорошая сводка по этому вопросу икеетсв в книге: Gates. The mutation factor in evolution. London. 1915. (стр. 349.)
7) Biolog. Centrbl. 36. 1916. (стр. 350.)