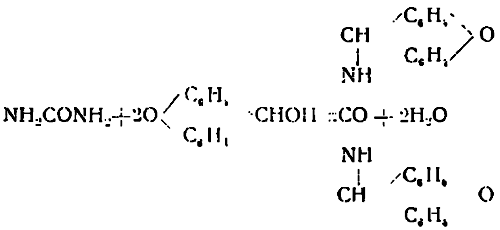
"Природа", №01-02, 1926 год, стр. 77-86.
Систематики растений и животных находят среди одноклеточных таких представителей, о которых приходится спорить, куда их отнести — к растениям или животным, настолько они являются похожими друг на друга. В этом нет ничего удивительного после того, что мы знаем в настоящее время об общности происхождения обоих отраслей органического мира. Сходство сохраняется и в том случае, когда мы от изучения морфологии и систематики переходим к разъяснению явлений жизни. Одной из важнейших задач физиологии является стремление дать обобщающую картину процессов, происходящих как в животной, так и в растительной клетке; поэтому всякая находка в растениях какого-либо вещества, которое было до сих пор известно только в животном организме, всегда дает ключ для понимания общности физиологических процессов.
Давно уже было подмечено различное отношение растений и животных к связанному азоту; в то время как первые с большим трудом черпают его из почвы, очень бережно его расходуют и никогда не выбрасывают, животныя в этом отношении являются как бы антиподами, т. к. все конечные продукты распада белка они выделяют из тела, как ненужные. Но, несмотря на это, в основных чертах превращение азота как у одних, так и у других — одинаково. Прежде всего надо отметить, что конечным продуктом превращения белков как у растений, так и у животных, является аммиак, который в свободном состоянии не может оставаться в организме, т. к. он является ядом для клетки. Как растения, так и животныя связывают его, причем обнаруживается одна интересная особенность — низшие растения (напр. плесневые грибы) и низшие животныя (актинии, пиявки) соединяют аммиак с вырабатываемой ими кислотой и в такой форме его обезвреживают, причем первые вновь пускают в ход аммиачную соль при синтезах белков, а вторые выделяют ее наружу. Иначе обстоит дело у высших животных и зеленых растений; здесь из аммиака при вторичном синтезе получаются амиды. При проростании семян, особенно в темноте, в них накопляется иногда очень большое количество амида аспарагина (CONH2 — CHNH2 — СН2 — СООН), а в других случаях глютамина (СООН — CHNH2 — СН2 — СН2 — CONH2), которые при выставлении проростков на свет опять используются для синтеза белка. У высших животных в печени из притекающего аммиака вырабатывается простой амид — мочевина (NH2CONH2), который через почки с мочей выбрасывается наружу.
Еще Буссенго, сравнивая мочевину животных с аспарагином растений. стремился дать обобщающую схему превращения азота, но только в последнее время мы получили новые данные, которые дополняют эту схему, благодаря нахождению мочевины в растительном мире.
В 1901 году Рейнке писал, что так как мочевина никогда не была найдена в растительной клетке, то, очевидно, что этот амид, происходящий при распаде белковых веществ, заменяется в растениях углекислым аммонием. Рейнке основывал свои суждения на нахождении в слизистых грибах углекислого аммония и на отсутствии там мочевины. Прошло всего два года после опубликования книги Рейнке, как австрийские химики Бамбергер и Ландзидль нашли в плодовых телах дождевиков (народное название гриба — "чортов табак") мочевину в количестве до 3,5% на сухой вес. Авторам присутствие мочевины казалось случайным. Они сперва думали, что мочевина попадает в грибы из мочи животных, находящихся на пастбищах, где дождевики хорошо произрастают. Но не найдя в грибах других элементов мочи и не открывши в самой почве мочевины, авторы пришли к выводу, что на нее (мочевину) надо смотреть как на продукт обмена этих грибов. После этого ряд ученых заинтересовался вопросом о нахождении и распространении мочевины в различных видах грибов. Сведения получились довольно противоречивые в одних и тех же грибах (напр., шампиньонах): то находили мочевину, то констатировали ее отсутствие.
Вопрос затруднялся тем обстоятельством, что прежние методы выделения мочевины в виде соединения ее или с щавелевой, или азотной кислотой, были неточные; трудно было выделить мочевину из смеси с другими веществами, когда она находилась в небольшом количестве в виду относительной растворимости указанных солей; если же количества мочевины невелики, то можно совсем не открыть мочевины там, где она заведомо присутствует. Поэтому большим приобретением для биохимии явился метод Фосса, лишенный всех вышеупомянутых недостатков. Метод заключается в получении кристаллического производного мочевины с ксантгидролом по уравнению:
Т. о. из одной молекулы мочевины и двух ксантгидрола получается одна молекула диксантил-мочевины, соединения, нерастворимого в воде и спирте. Этот метод дает возможность производить осаждение мочевины из сильно загрязненной посторонними примесями среды. Полученное в этой реакции соединение обладает высоким молекулярным весом — 420, в то время как мочевина имеет только — 60; т. е. производного продукта получается в 7 раз больше, чем было по весу мочевины; последнее обстоятельство приобретает особенно важное значение при определении очень малых количеств мочевины; удается производить учет 2—3 мгр. мочевины, а микроскопически распознать даже 0,01 мгр., т. к. диксантил-мочевина кристаллизуется в виде характерных шелковистых иголочек. Метод дает возможность открыть мочевину при разбавлении 1:1.000.000. Благодаря изумительной чувствительности метод Фосса должен получить широкое практическое применение; известно, что при некоторых заболеваниях в плазме крови появляется некоторое количество мочевины; это можно учесть в небольшой пробе крови, пользуясь ксантгидрольным методом. Прежние способы, конечно, для таких целей не годились.
Я применил этот метод для выяснения значения мочевины в растительном мире. Еще в 1916 году, работая с дождевиками, я пришел к выводу, что количество мочевины в плодовых телах гриба, снятых с мицелия, возрастает по время созревания, а в спорах ее меньше, чем в капиллиции. С первых же опытов выяснилось, что дождевики являются в высшей степени подходящим материалом для выяснения вопроса о превращении мочевины. Так, в одном опыте было взято две партии мелких незрелых плодовых тел дождевика (Lycoperdon piriforme): одна (А) была высушена немедленно, обращена в порошок и доведена до постоянного веса, другая (В) оставалась в течение 7 дней на бумаге при обыкновенной температуре; грибы за это время дозрели, в них образовались споры, порошок из них получился темнозеленый. За время созревания (В) количество общего азота в этой порции возрасло в виду потери большого количества углеводов в процессе дыхания, а вместе с этим увеличилось и процентное содержание в них мочевины:
| Колич. N в %% |
Мочевина в мгр. |
|||
| Порция | А | 5,75 | 18,3 | |
| "" | В | 6,92 | 84,2 |
Такое увеличение мочевины при созревании замечалось при всех опытах. В других случаях плодовые тела разрезались вдоль на две равные части: в одних половинках мочевина определялась сразу, в других (опытных) после 3—5-дневного созревания; так, в одном опыте в контрольных половинках оказалось 83,6 мгр. мочевины, а в опытных — 136,1 мгр. Мои наблюдения показали, что накопленная при созревании мочевина, затем начинает исчезать в спорах, где ее количество опускается до нуля.
В дальнейшем я пользовался плодовыми телами дождевиков, взятых с одного и того же мицелия, но в различные стадии зрелости. В следующем опыте были взяты четыре стадии зрелости (I—IV) дождевика (Lycoperdon piriforme): I — самая молодая, IV — вполне зрелая. Из таблицы видно, что количество мочевины постепенно наростает, а затем падает в зрелых экземплярах:
| Стадии зрелости: | % мочевины на сухой вес. |
|
| I | 0 | |
| II | 1,1 | |
| III | 4,6 | |
| IV | 0,07 |
Я проделал большое количество определений в различных грибах, находящихся на различных стадиях зрелости, и обнаружил, что в некоторых из дождевиков количество мочевины доходит до 11,16% (Bovista nigriscens) на сухой вес, при чем содержание зависит от вида дождевика; так, найдено мочевины у:
| Lycoperdon | saccaturn | ..... | 2,85 | % | на | сухой | вес. |
| " | piriforme | ..... | 4,62 | " | " | " | |
| " | gemmatum | ..... | 10,7 | " | " | " | |
| " | molle | ..... | 9,2 | " | " | " | |
| " | marginatum | ..... | 5,8 | " | " | " | |
| " | echinatum | ..... | 1,16 | " | " | " |
Необходимо подчеркнуть, что это наивысшее наблюденное количество мочевины; в более молодой и зрелой стадии количество ее может спуститься до нуля; кроме того, никак нельзя поручиться, что мои цифры суть наивысшие для каждого вида гриба; очень может быть, что я не уловил момента, когда мочевины накопилось наибольшее количество. Таким образом, удалось показать огромное накопление мочевины в некоторых грибах, при чем в отдельных случаях оказалось, что около половины всего азота гриба приходится на мочевинный.
В дальнейшем было обнаружено, что даже один и тот же вид варьирует в содержании мочевины в зависимости не только от стадии зрелости, но и от питания азотом, которое сказывается на процентном содержании азота в плодовом теле. Так, вот какие соотношения наблюдались в ряде случаев у выращенных в различных условиях азотистого питания экземпляров Lycoperdon piriforme:
| %-е содержание азота. |
Наивысшее наблюденное %-е содержание мочевины, считая на сухой вес. |
|
| I | 7,92 | 4,62 |
| II | 5,75 | 0,55 |
| III | 4,19 | 0 |
Отсюда ясно, что при малом доступе азотистого питания (III случай) даже в стадии перед созреванием мочевины не оказалось.
Интересно, что одна и та же грибница культурных шампиньонов (Psalliota campestris), пророставшая в несколько отличных условиях азотистого питания, дала при развитии плодовых тел в двух случаях такие колебания в содержании мочевины:
| I — от 1,48 до 2,86% | мочевины |
| II — от 3,47 до 4,12% |
Лучшие условия питания сказались в увеличении мочевины, а когда во втором случае азотистое питание было еще улучшено внесением его в почву, то цифра мочевины еще возрасла до 5,6%; в данном случае ясно видна зависимость между содержанием мочевины и питательной средой, на котором грибы живут.
Всеми авторами принимается, что мочевина в животном организме образуется путем ферментативного синтеза из аммиака (NH3) и углекислоты и что фермент действует в водной среде. Относительно образования мочевины в растении Горис и Маскре допускали, что она возникает в них во время высушивания и что в некоторых грибах существуют азотистые вещества, которые дают в числе продуктов распада мочевину. В животной физиологии хорошо известно, что мочевина может образоваться из продуктов гидролиза белка, именно из аргинина:

т.-е.: аргинин + вода дают мочевину + оринтин. Растения также могут образовывать мочевину из аргинина; в последнем нас убеждает широкое распространение фермента аргиназы, как это показано работами А. Р. Кизеля; но количества мочевины из аргинина не могут быть большими и во всяком случае не могут покрыть тех больших ее запасов, которые накапливаются в грибах и доходят до 11,6% на сухой вес.
В моих опытах было показано, что мочевина в грибах образуется, при созревании их, и что это совпадает с сильным окислительным процессом, во время которого тратится большое количество без-азотистого материала. Зная, что мочевина может возникать из аммиака и углекислоты, я стал давать созревающим дождевикам аммиак и следил, не образуется ли за счет его мочевина; все мои опыты дали положительный результат. Я брал плодовые тела дождевиков или их половинки, и одну порцию оставлял контрольной, а другую предварительно в течение нескольких часов или обрабатывал газообразным аммиаком, или давал его в растворе в виде аммонийной соли; обе порции оставлялись в течение 3—4 дней дозревать, а затем высушивались, превращались в порошки и шли на определение мочевины.
Так, в одном опыте получилось в половинках:
| Мочев. в мгр. | % мочевины на сухой вес. |
||
| Контрольная | ...... | 135,4 | 2,08 |
| Опытная | ...... | 191,2 | 2,81 |
т. е. в порции, обработанной аммиаком, мочевины оказалось на 41% больше, чем в контрольной.
В другом ряде опытов удалось показать, что накопление мочевины происходит и в тех случаях, когда плодовые тела дождевиков не были сняты с грибницы, на которой они росли; аммонийная соль в данном случае в растворе вводилась в почву, и было констатировано, что поглощенная аммонийная соль переходила в мочевину. Судьба аммиака в данном случае вполне аналогична тому, что мы наблюдаем в печени высших животных, где также из аммиака и углекислоты формируется мочевина. Упомянутые опыты еще раз указывают, что образование мочевины идет в живых грибах и есть важный физиологический процесс растения.
Широкое распространение мочевины в растениях и большое ее накопление в отдельных случаях ставит перед нами вопрос о физиологическом ее значении. После того, как мною было показано, что растительная клетка может самостоятельно готовить мочевину, должны в значительной мере отпасть утверждения Фосса, что мочевина вполне или отчасти черпается из почвы, на которой растения произрастают. Некоторое, очень малое количество мочевины имеется в почве, и нет ничего удивительного, что она оттуда всасывается, тем более, что мочевина легко проникает в растительную клетку. Мои новые опыты показывают, что если плодовое тело шампиньона погрузить пеньком в 1% раствор мочевины, то в шляпке количество мочевины увеличивается с 3,0% до 14,9% на сухой вес; особенно интересно, что вся масса мочевины направляется в пластинки (гимениальный слой) гриба, где и откладывается; так, о одном опыте с всасыванием мочевины в шампиньоне ее оказалось:
| В части шляпки без пластинок | ...... | 5,7% |
| В пластинках | ...... | 13,4% |
Допуская в естественных условиях некоторое проникание мочевины из почвы, мы должны признать, что главная ее масса приурочивается к важнейшему физиологическому процессу — плодоношению; к моменту созревания количество мочевины наибольшее; затем количество ее уменьшается и она совершенно исчезает в спорах. Какое же значение мочевины? Мы знаем, что высшие растения не накапливают в себе аммиака; образующийся в результате расщепления белковых веществ аммиак чаще всего отлагается в растении в виде амидов — аспарагина и глютамина. Более того, опыты японцев Киношита и Сузуки показали, что при питании растений в водных культурах аммиачными солями за их счет образуется аспарагин. Особенно показательными в этом отношении являются исследования Д. Н. Прянишникова и его учеников, которым удалось удвоить количество аспарагина при питании растений хлористым аммонием. Эти данные вполне могут быть поставлены в сравнение с явлением накопления мочевины в грибах в момент спорообразования за счет введенного извне аммиака; в этот момент происходит интенсивная окислительная работа, а давнишние опыты Палладина представили перед нами процесс образования аспарагина, как окислительный. Накопление аспарагина — временное явление, ибо при доступе углеводов он опять используется растением на образование белковых веществ. То же самое наблюдается у грибов с превращением мочевины, которая в случае надобности пускается растением в круговорот азотистого питания. Все эти факты приводят меня к убеждению признать за мочевиной, накопляющейся в дождевиках, запасное азотистое вещество, аналогичное амидам зеленых растений — аспарагину и глютамину.
Все опыты с созреванием дождевиков указывают, что мочевина исчезает в спорах, но это не значит, что образовавшийся благодаря ее разложению аммиак там накопляется; напротив, последний не удается констатировать; отсюда мы делаем вывод, что аммиак в дальнейшем переходит в сложные азотистые нерастворимые соединения.
Плодовые тела дождевиков являются очень удобным объектом для изучения превращения мочевины, т. к. главную массу их составляют споры, количество которых достигает 75% на вес плодового тела. Другая картина должна получиться, если мы от дождевиков обратимся к таким содержащим мочевину шляпным грибам, у которых количество спор ничтожно, сравнительно с большим весом всего плодового тела. Эти соображения руководили мною, когда я решил исследовать превращение мочевины у шампиньонов. Эти грибы представляют то удобство, что разводятся в теплице и растут круглый год. Количество мочевины у шампиньонов зависит от азотистого питания, а также и от возраста; когда я в одном случае проанализировал четыре стадии грибов, выросших из одной и той же грибницы, то у меня получились такие данные:
| Стадии: | Мочевина в % на сухой вес. |
|||
| I | — | самая молодая | ...... | 2,6% |
| II | — | молодая | ...... | 3,4% |
| III | — | начинающая созревать | ...... | 5,7% |
| IV | — | зрелая | ...... | 12,4% |
Здесь выяснилось, что у зрелых старых грибов количество мочевины не только не уменьшилось, а наоборот, достигло рекордной величины 12,4%, а в другом случае даже 13,2% на сухой вес. Но зато в собранных у шампиньонов спорах количество мочевины опять сильно упало и достигло 1,1%.
В дальнейшем удалось показать, что при созревании шампиньонов идут автолитические процессы, при которых белковые вещества, а, вероятно, и хитин, расщепляются; из продуктов их распада при окислительном синтезе образуется мочевина. В опытах нз шляпки шампиньона бралась проба (вырезалась ¼ часть), которая немедленно высушивалась, а оставшаяся доля ставилась пеньком в воду на 2—3 дня, а затем шляпка этой опытной порции также высушивалась и шла на определение мочевины. В одном опыте было в шляпке 9,7%, а стало через 2 дня 12,9% мочевины, но наряду с этим было констатировано уменьшение нерастворимого в воде азота; в этом случае всего азота в грибе было 134,8 мгр., а мочевинного азота оказалось 63,9 мгр., т. е. 47,4% всего азота приходится на мочевинный. Получается впечатление мочевинного отравления гриба.
Такие большие количества мочевины накапливались в шампиньонах, растущих в искусственной культуре при огромном избытке азотистого питания. Плодовое тело получает азот из почвы и накопляет его сначала в виде сложных соединений, затем к моменту формирования спор эти вещества разлагаются и, проходя стадию аминокислот, переходят в аммиак, который только в небольшой части используется в спорах, а в главной массе переходит в мочевину, которая и остается в шляпке гриба до конца его жизни. Гриб умирает обогащенный мочевиной. Интересно, что образование мочевины связано с нормальным окислительным процессом дыхания; если часть шампиньона оставить под стеклянным колпаком, где имеются пары хлороформа, то автолитические процессы в тканях гриба будут итти и белок будет расщепляться до аминокислот, но превращения последних в мочевину мы не будем наблюдать.
Очевидно, эти условия автолиза гриба не тождественны с теми, которые идут в живом организме. Т. о. в шампиньонах мочевина уже не играет роли запасного вещества1).
Мне удалось показать, что и в чистой культуре на желатине с бражкой в мицелии шампиньона также накопляется мочевина; в одном опыте в такой культуре гриба, выросшего в пробирке, после 5 месяцев удалось обнаружить мочевину: азота в пленке шампиньона оказалось — 4,0 мгр., а мочевины в культурной среде — 5,4 мгр.
Интересно, что в данном случае мочевина не распалась, несмотря на длительный срок хранения культуры. Это зависит от отсутствия в данный момент в шампиньонах уреазы фермента, расщепляющего мочевину по уравнению: СО(NH2)2 + 2Н2O = СО2 + 2NH3.
Действительно, все попытки экспериментально доказать присутствие уреазы в шампиньонах и дождевиках не удаются, потому что ткани этих грибов, оставленные с мочевиной, не разлагают последнюю с выделением аммиака. Но мы должны допустить, что в этих живых грибах должна существовать уреаза; иначе было бы непонятно, как может использоваться мочевина в спорах дождевиков, потому, что если бы там не было уреазы, то мочевина явилась бы продуктом отброса и растение должно бы было его выбрасывать, так же, как и животное.
Теперь мы имеем уже довольно большой список грибов, где встречается мочевина; но в последнее время удалось показать, что она имеется не только в грибах, но и в других растениях, но только в следах; так, Фосс выдавил сок из 10 килограммов листьев цикории и получил своим методом небольшое количество кристаллов производного мочевины, давшее ему возможность определить точку плавления; небольшие количества мочевины были найдены автором и у тыквы, дыни, цветной капусты, брюквы, шпината, моркови и картофеля; но вряд ли это нахождение имеет большое физиологическое значение. Правда, некоторые авторы, напр. Уернер, считают, что всегда при конечном распаде белков должна образоваться мочевина; в развитых растениях мочевина встречается редко и случайно, потому что там идет белковый синтез, но этот взгляд пока не доказан. В дальнейшем стало ясно, что мочевина в грибах появляется в результате избыточного азотистого питания и что шампиньоны в чистой культуре то образуют ее, то нет в зависимости от того — дадим ли мы им много или мало белка в среду, на которой они живут. Оказалось, что избыток углеводного питания также задерживал образование мочевины. Все опыты со стерильной культурой шампиньонов протекают крайне медленно; нужно ждать месяцы для получения результатов, поэтому в самое последнее время я обратился к выращиванню плесневых грибов, которые развиваются в термостате в стерильных условиях уже через 5—10 дней. Но для достижения своей цели я изгнал из культурной жидкости углеводы, а давал, кроме питательных солей, в каждом отдельном случае по 3 гр. пептона, который и являлся для гриба, как азотистым, так и углеродистым материалом. В одном опыте на 100 к. с. питательной жидкости обычный плесневой гриб аспергилл развил через 12 дней 48,6 мгр. мочевины, а пленка гриба в сухом виде весила 0,63 гр., т. е. мочевины получилось 7,5%, считая на вес гриба.
Было обнаружено, что вся мочевина содержалась в питательной жидкости, на которой гриб развивался, т. к. в нем самом мочевина отсутствовала. Гриб, выросши в отсутствии углеводов, был нацело лишен уреазы, фермента расщепляющего мочевину, между тем как давно было известно, что при обычной культуре на сахаре аспергилл всегда этот фермент содержит. Таким образом, мочевина в нем явилась отбросом. Но понятие отброса для растений не является однозначущим с тем, что принимается для животных. Например, для последних углекислота всегда отброс, выводимый при дыхании из организма, но для зеленого растения углекислота в присутствии солнечного света — питательное вещество и только в темноте является как и для животных, отбросом. Нечто подобное мы наблюдаем и с мочевиной в грибах; она накапливается и не может потребляться, если нет углеводов, но как только эти последние даются, мочевина опять пускается в оборот как питательное вещество. Это можно хорошо видеть в опытах над плесневыми грибами выросшими на пептоне или желатине; если в такую питательную жидкость, в которой накопилась мочевина, прибавить сахар, то гриб начнет быстро расти и мочевина, как и сахар, исчезнет из питательной среды; пленка же гриба теперь будет содержать богатую уреазу, которой не было до прибавления сахара. Из этих опытов вытекает, что один и тот же гриб может то содержать мочевину и быть лишенным уреазы, то наоборот — все будет зависеть от питательной среды.
Вся образующаяся в этих опытах при развитии плесневых грибов мочевина происходит из аргинина, содержащегося в пептоне и желатине. Действительно, мочевина может получиться при расщеплении аргинина на орнитин и мочевину, а эта последняя будет накапливаться в виду отсутствия уреазы.
Новые данные о мочевине дают нам возможность представить более обобщающую картину превращения азота у растений и животных. В животном и растительном организмах конечным продуктом распада белка является аммиак, который при окислительном синтезе в печени животных переходит в мочевину, а у зеленых растений при недостатке углеводов переходит в амиды — аспарагин и глютамин; грибы же в этом отношении оказываются стоящими ближе к животным, т. к. у них из аммиака синтезируется мочевина, то как запасной продукт (у дождевиков), то отчасти как запасной, то как продукт отброса (у шампиньонов и плесневых грибов). Последний случай дает нам возможность наблюдать небывалое явление в растительном мире — перевод азота в отброс в виде мочевины. Чем больше мы изучаем растительную и животную клетку, тем больше мы находим фактов, которые дают нам возможность сблизить их не только морфологически, но и физиологически. Добытые факты над превращением мочевины в растительной клетке являются в этом вопросе хорошей иллюстрацией.
1) Но это количество мочевины, которое направляется в споры и там потребляется, должно быть также названо запасным, как у дождевиков, и не потребляется далее, а имеет характер отброса, который формируется из вредного для клетки аммиака, как в печени высших животных, и остается в такой нейтрализованной форме до конца жизни.
(стр. 84.)